目
| 目(眼) | |
|---|---|
 ヒトの左眼 | |
 1:硝子体 2:鋸状縁 3:毛様体筋 4:チン小帯 5:シュレム管 6:瞳孔 7:前眼房 8:角膜 9:虹彩 10:水晶体皮質 11:水晶体核 12:毛様体突起 13:結膜 14:下斜筋 15:下直筋 16:内側直筋 17:網膜の動静脈 18:視神経乳頭(視神経円板) 19:硬膜 20:網膜中心動脈 21:網膜中心静脈 22:視神経 23:渦静脈 24:テノン嚢 25:黄斑 26:中心窩 27:強膜 28:脈絡膜 29:上直筋 30:網膜 | |
| ラテン語 | Oculus |
| 英語 | Eye |
| 器官 | 感覚器 |
| 動脈 | 眼動脈 |
| 静脈 | 眼静脈 |
| 神経 | 視神経 |
目(眼、め)は、光を受容する感覚器である。光の情報は眼で受容され、中枢神経系の働きによって視覚が生じる。
ヒトの眼は感覚器系に当たる眼球と附属器[1]、神経系に当たる視神経と動眼神経からなる[2]。眼球は光受容に関連する。角膜、瞳孔、水晶体などの構造は、光学的役割を果たす。網膜において光は神経信号に符号化される。視神経は、網膜からの神経情報を脳へと伝達する。付属器のうち眼瞼や涙器は眼球を保護する。外眼筋は眼球運動に寄与する。多くの動物が眼に相当する器官を持つ。動物の眼には、人間の眼と構造や機能が大きく異なるものがある。
以下では、まず前半でヒトの眼について、後半では動物全体の眼についてそれぞれ記述する。
目次
1 ヒトの眼の構造
1.1 眼球
1.2 付属器
1.3 眼の血管系
1.4 眼の求心性神経
1.5 眼の遠心性神経と筋
2 眼の発生
2.1 水晶体と硝子体
2.2 視神経と網膜中心動静脈
2.3 網膜
2.4 強膜、脈絡膜、毛様体
2.5 角膜
2.6 虹彩
2.7 外眼筋
3 眼の機能
3.1 眼光学系
3.2 網膜
3.3 付属器
3.4 コミュニケーション機能
4 眼の疾患
5 動物の眼
5.1 光受容器
5.2 散在性視覚器
5.3 眼点
5.4 杯状眼
5.5 窩状眼
5.6 水晶体眼
6 目の形態
6.1 単眼
6.2 複眼
6.3 カメラ眼
7 水棲と陸棲
8 眼の配置
9 視覚
9.1 視力
9.2 色覚
10 眼の進化
11 脚注
12 参考文献
13 関連項目
14 外部リンク
ヒトの眼の構造

ヒトの眼の構造
眼は眼窩に位置し、眼球、視神経、付属器からなる[1]。
眼球
眼球は外壁(眼球壁)と内容物からなる。外壁は3層構造をなし、最外層で血管が少ないため白く見える外膜(繊維膜)は前部1/6に当たる強く湾曲した屈折を担う角膜と[3]その外周にあり[3]5/6を覆う強膜からなる[1]。
中間層は眼球血管膜(俗称:ぶどう膜[3])と総称され、カメラの絞りに相当する膜状組織であり中央に瞳孔と呼ばれる開口部を持つ[3]虹彩[3]、中に筋肉を持ち水晶体を支えたりひっぱたりする毛様体[3]、メラニン色素を多く含み光の乱反射を防ぐ脈絡膜からなる[1]。
最内層は神経性網膜と色素上皮からなる網膜がある。網膜は、後半部に受けた光を神経の興奮状態に替えて視神経に伝える役割を持つ網膜視部と、前半部の光を感じない網膜盲部がある。この境界には鋸状縁という。また、眼球奥にも視神経に繋がるためくぼんだ箇所(視神経円板・視神経乳頭)があり、光を感じない。この場所から約4mm外側には中心窩と呼ばれる窪みがある。ここは視力が最も高く発揮される場所であり、物を凝視する際の焦点として使われる[1]。
眼球の内容物には、直径約1cmの弾性を持つ凸レンズ状組織である水晶体、無色透明なゼリー状で眼球の形状を支える硝子体、眼圧を調整する液体の眼房水がある。角膜と虹彩のあいだを前眼房、虹彩と水晶体・硝子体のあいだを後眼房とよぶ[1]。眼房水は毛様体の上皮で分泌供給され、強膜静脈洞(シュレム管)から排出されて静脈に還流する[1]。
視神経は網膜神経節細胞の軸索である。視神経は視神経乳頭から視交差へ向かう。視神経は視交差以降では視索となり外側膝状体、上丘、視交叉上核などへ向かう。視神経は視神経鞘に包まれる。視神経鞘には網膜中心動脈、網膜中心静脈なども包まれる。
付属器
眼球の外側には付属器が付く。眼瞼は眼球の前方に位置する折りたたみ可能な上下2枚の襞で、眼球の保護をする。内側は血管と神経が張り巡らされた結膜があり、この箇所は感染症を起こしやすい。内部には眼輪筋と、結合組織からなり眼球と瞼の摩擦を低減する脂肪性物を分泌するマイポーブ腺を一列に備えた瞼板がある[1]。
涙器には涙腺と多数の涙道がある。平時には少量の涙を分泌し眼球の乾燥を防ぎながら、時には多量の涙を流すこともある。通常では涙は目頭の方へ集まり、涙小管から鼻根部にある涙嚢そして鼻涙管を通じて鼻腔に落ちるようになっている[1]。
筋肉(眼筋)は眼窩の中にあり眼球を動かす横紋筋である。6本のうち上直筋・下直筋・内側直筋・外側直筋の4本は直筋群と呼ばれ、眼球前方の強膜と連結して眼球をそれぞれが収縮する方向に向ける働きを持つ。残りの上斜筋・下斜筋は同じく眼球をある方向に向ける役割を担うが、その末端は滑車と呼ばれる腱と繋がっている[1]。
眼の血管系

網膜中心動脈と網膜中心静脈は眼底写真により観察できる。
眼を栄養する主要な動脈は、内頸動脈の枝である眼動脈の分枝である。網膜中心動脈は視神経管を通り、網膜の内面に分布する。短後毛様体動脈は視神経の近くで強膜を貫いて脈絡膜に分布する。
眼瞼や外眼筋には眼動脈の分枝のほかに、外頸動脈の枝である顎動脈、浅側頭動脈、顔面動脈などの分枝も分布する。
眼の主要な静脈は眼静脈である。眼静脈は上眼静脈と下眼静脈を受ける。網膜中心静脈は網膜中心動脈を受け、上眼静脈へ注ぐ。脈絡膜や毛様体に分布した血液は渦静脈となり、上眼静脈へ注ぐ。眼静脈は海綿静脈洞へと注ぐ。
網膜の特に後方奥は眼底と言い、瞳孔を通じて検視鏡で観察できる。ここは、人体において直接に血管を見ることが出来る唯一の場所であり、動脈硬化など血管の診断に利用される[1]。
眼の求心性神経
視神経は網膜神経節細胞の軸索である。視神経は視神経溝で視交叉を作り、外側膝状体や上丘へと投射する。
三叉神経第1枝の眼神経の枝である、涙腺神経、眼窩上神経と滑車上神経からなる前頭神経、滑車下神経は、眼瞼や角膜の知覚に寄与する。長毛様体神経は毛様体と虹彩の知覚に寄与する。
眼の遠心性神経と筋

外眼筋は眼球に付着し、眼球運動に寄与する。
眼瞼の筋のうち、上眼瞼挙筋は動眼神経支配、眼輪筋は顔面神経支配の横紋筋である。瞼板筋は交感神経支配の平滑筋である。それぞれの筋は眼瞼の運動に関与する。眼瞼の運動には前頭筋も関与する。
内眼筋は毛様筋と虹彩筋からなる。毛様体筋は動眼神経支配であり、毛様体神経節からの短毛様体神経により支配される。虹彩筋のうち瞳孔括約筋は動眼神経支配、瞳孔散大筋は交感神経支配であり、瞳孔径の変化に寄与する。
下斜筋を除く外眼筋は、総腱輪を起始として強膜に停止する。下斜筋は眼窩前部底から起始する。上直筋、下直筋、内直筋、下斜筋は動眼神経核から出る動眼神経支配である。上斜筋は滑車神経支配である。外直筋は外転神経支配である。
動眼神経のうち動眼神経副核由来のものは副交感神経性であり、毛様体神経節を経て短毛様体神経となり、毛様体筋と瞳孔括約筋に分布する。
三叉神経第2枝の上顎神経からは、翼口蓋神経節を経て涙腺へ分布する副交感性の枝が出る。
眼の発生

眼胞は脳室と連続する。眼胞の二重壁は、色素性網膜と神経性網膜になる。外胚葉の一部は陥凹して水晶体窩となる。
眼を構成する組織のうち、角膜の外層と水晶体は外胚葉由来である。網膜、視神経は神経外胚葉由来である。強膜、脈絡膜、毛様体、角膜の内層は中胚葉および神経堤由来である。
眼原器は胎生第2週に、神経管の前脳から形成される。前脳のこの領域は、将来の間脳となる。眼原器は外側に拡張し、第3週に眼胞となる。眼胞の内部は、将来の第三脳室と連絡する。眼胞は周囲を間葉組織に取り囲まれ、将来の視神経鞘となる。第4週には、眼胞の先端部が陥凹し、二重壁を持つ眼杯と、二重壁を持たない眼茎となる。

水晶体窩は最終的に表層から完全に分離して水晶体胞となる。
水晶体と硝子体
眼杯の前面に位置する外胚葉が陥凹して水晶体窩となる。水晶体窩は最終的に表層から完全に分離して水晶体胞となる。水晶体胞の後面と眼杯の前面とのあいだには、一次硝子体が形成される。水晶体胞の後面には一次水晶体線維が形成される。一次硝子体は、二次硝子体が生じると退縮する。
視神経と網膜中心動静脈

網膜中心動脈は視神経に取り込まれる。
眼杯の腹側面と眼茎に溝ができ、これがつながって眼杯裂となる。眼杯裂は間葉と眼杯を連絡する。眼杯裂には硝子体動静脈が含まれる。硝子体動脈は眼動脈の枝であり、水晶体や眼杯の内部の組織を栄養する。硝子体動脈はその後、網膜以外への枝を失い、網膜中心動脈となる。眼杯裂はその後、眼杯と眼茎に取り込まれる。最終的には視神経の軸索に取り囲まれる。
網膜
眼杯の内壁では神経細胞への分化が生じ、神経性網膜となる。外壁はメラニン色素を含む網膜色素上皮となる。内壁と外壁はもともとは第三脳室と通じる腔により隔てられていたが、徐々に腔は狭小化し、最終的に消失する。眼杯の内壁は視細胞などの細胞へと分化する。神経節細胞の軸索は、眼茎の壁内を進む。線維が増大するにつれて眼茎の内腔は狭小化して、内腔のない視神経となる。
強膜、脈絡膜、毛様体

水晶体胞の周囲には間葉組織が侵入する。
間葉組織は眼杯を取り囲み、2層の構造を生じる。外層は強膜、内層は脈絡膜を生じる。強膜は硬膜と、脈絡膜は軟膜とクモ膜と相同である。
水晶体が形成されると、眼胞を取り囲んでいた間葉組織は、水晶体と体表との間に侵入する。後にこの部位の間葉組織は、前後の2層に分かれる。前層は角膜固有質となり、角膜の内層となる。後層を前眼房の中皮とよぶ。両者の間は将来の前眼房となる。前眼房の中皮の水晶体に接する部分は後に崩壊して、水晶体の間に間隙を生じる。この間隙は将来の後眼房となる。前眼房の中皮の残存層は瞳孔膜となり、後に穿孔して瞳孔を生じる。
毛様体は眼杯と周辺の脈絡膜により生じる。毛様体色素上皮は網膜色素上皮と連続する。毛様体無色素上皮は神経性網膜と連続するが、神経細胞は存在しない。
角膜
角膜の内層は前眼房の中皮に由来し、中胚葉由来である。角膜の外層は体表外胚葉に由来する。
虹彩
眼杯の前縁とその周囲の間葉は眼杯の内側へ進出して、水晶体を部分的に覆う。このことにより、虹彩を生じる。虹彩筋は、神経堤由来の平滑筋である。虹彩の血管や結合組織は間葉由来である。
外眼筋
外眼筋は間葉から生じる。上眼瞼挙筋は上直筋から分かれて生じる。
眼の機能
眼光学系

角膜と水晶体は光を屈折させる。
光は角膜により屈折される。次に瞳孔を通過する。次に水晶体により屈折される。最後に網膜へと投射する。
角膜の屈折力は40 D程度である。これは、水晶体の屈折力20-30 Dよりも大きい。角膜の屈折力が大きいのは、角膜が屈折率の差の大きい空気と境界をなしているためである。角膜の屈折力は大きいため、角膜の障害により視力は大きく低下する。
瞳孔径は2-8mm程度の間で変化する。瞳孔径の変化は虹彩をなす虹彩筋の作用による。瞳孔径の変化は光量に依存する。虹彩の色はメラニン色素により決まり、個体差がある(→ヒトの虹彩の色)。
水晶体の屈折力は20-30 D程度のあいだで変化する。水晶体は、毛様体筋の働きによって厚みを変化し、屈折力が変化する。この作用を調節とよぶ。調節機構の説明としては毛様体小帯の緊張により、水晶体の周囲が圧迫されて調節されるとする緊張説と毛様体筋の収縮により毛様体小帯が弛緩し水晶体が球形に近づくことで起こるとする弛緩説とに分かれる。[4]
眼光学系は、カメラにたとえられることがある。角膜は単焦点レンズ、瞳孔は絞り、水晶体は可変焦点レンズ、網膜はフィルムに相当するとされる。しかしながら、眼光学系の各々の屈折面では明確な光軸は定義されない。また、各々の屈折面における近似的な光軸は、互いに一致しない。さらに、中心窩や瞳孔の中心は、近似的な光軸上には位置しない。このように、眼光学系はカメラのような共軸光学系とは異なり、非共軸光学系である。
網膜
網膜において光受容がなされる。すなわち、眼光学系を通じて網膜に投射された光は、網膜において神経信号へ符号化される。網膜からは視神経が出て、神経信号を外側膝状体や上丘へと伝達する。

涙器は涙液を分泌して角膜を保護する。
付属器
眼瞼は眼球を物理的に保護する。また、瞬目により結膜表面に涙液を分布させる。
涙器は涙液の分泌と除去に関わる。涙液にはリゾチーム、ラクトフェリン、免疫グロブリンなどが含まれる。
コミュニケーション機能
少なくとも人間の場合に、眼は表情を構成する重要な要素であり、視覚的な個体間の情報交換、いわゆる非言語コミュニケーションの大きな部分を担っている。日本語では「眼は心の窓」「目は口ほどにものを言い」という表現がある。また、「眼が泳ぐ」「目が据わる」などの表現も、眼の感情表現における役割を示すものである。目と目を合わせることをアイコンタクトと言う。
さらに、ヒトの場合はまぶたの間から虹彩より外の白目の部分が見えること、その上に眉毛があることは独自の特徴で、これらは眼の作る表現の幅を広げ、強調する役割を担ってもいる。また涙も単に眼を洗浄する役割以上に感情と強い結びつきを持つ。
眼の疾患
水晶体が濁って起こる白内障や網膜神経節細胞が死滅する緑内障、網膜から神経網膜が剥がれる網膜剥離、結膜にできる炎症である結膜炎など、目を患部とする疾患は数多く存在する。特に眼球部分を患部とする疾患の場合、症状の進行によって失明することもある。目を扱う医学の診療・研究分野は眼科学と呼ばれる。また、目において屈折異常が起きると近視や乱視、遠視などといった症状が現れる。こうした屈折異常を根本的に治療することは困難であるが、眼鏡やコンタクトレンズといった矯正器具を使用することでほとんどの屈折異常は生活上問題のない程度まで症状を緩和することが可能である。
眉毛と目の組み合わせを左に90度回転させると「10」に見えることから、毎年10月10日は「目の愛護デー」とされており、眼科医などが目の異常などの早期受診などを呼びかける啓発行事を実施している[5]。同様な理由で、1001=10月1日は「眼鏡の日」となっている。
動物の眼
動物の眼は、発生起源が皮膚の表層部である表皮であるものと、中枢神経系である脳の一部から生じるものの2つに大別できる。無脊椎動物の眼は皮膚由来であり、脊椎動物の眼は脳由来である[6]。
光受容器
原生生物のミドリムシは、鞭毛基部に感光部を持つ。多細胞生物のうち、光に応答するが、光を受容するための特別の構造を有さない動物は、体表の細胞に感光性色素を持つ。これらの構造は、光受容のために分化した構造ではない。
散在性視覚器
最も原始的な眼は環形動物であるミミズのような明暗を感知するだけの「明暗視」が可能なものが体表面に分布する形態のものである。このような体の各部に分布する眼は「散在性視覚器」(さんざいせいしかくき)と呼ばれ、ミミズでは表皮の表皮細胞の間に単独の視細胞がまばらに分布している[6]。ミミズの光受容性表皮細胞は、例外的に光受容膜に類似した微絨毛を持つ。
眼点

貝殻の端に並ぶ黒い点が、ホタテガイの眼点である。約80個存在する。
視細胞が集まったものが「眼点」(がんてん)である。眼点は体表の視細胞が色素を含んだ支持細胞に裏打ちされることで「網膜」を形成するが明暗視だけが可能であり光の方向は判別できない。腔腸動物のクラゲや扁形動物のプラナリアなどが眼点を持つ。プラナリアや軟体動物のホタテガイなどごく少数の無脊椎動物では視神経が光の入射側になる「背向性眼」であり、これらは無脊椎動物の眼としては例外的である[6]。
杯状眼
視細胞と支持細胞から成る網膜組織が体表面から陥凹し、杯状となりレンズを備えない構造を形成したものが「杯状眼」(はいじょうがん)である。杯状になることで光の入射方向を判別できる。腹足類のカタツムリ[6]やカサガイなどが杯状眼を持つ。
窩状眼
杯状眼では光の入射方向の判別精度があまり高くなく、杯の入口が狭くなって穴となったものが「窩状眼」(かじょうがん)である。杯状眼でもコロイド状分泌物で凹部を満たすものもあるが、窩状眼でも穴となった内部に硝子体を備えることで性能を高めるようになっている[6]。
水晶体眼
窩状眼では網膜へ光線を絞って投影できるが、光の入射口が狭く多くの光を無駄にすることになる。光を屈折させる専用のレンズである水晶体を備え、前面にはそれを保護する角膜を持ったものが「水晶体眼」である。無脊椎動物としてはホラガイのような貝類から水晶体眼を持つようになり、脊椎動物の眼も水晶体眼である。イカやタコといった頭足類の眼は無脊椎動物の中でも特に発達した水晶体眼であり、脊椎動物の眼と形態がかなり似る[7]までに進化を遂げているが、発生過程では眼の組織が表皮から生じるため、眼胞と呼ばれる眼の組織が表皮細胞から分化して生じてその前壁部分が厚みを帯びて水晶体後半となり、表皮細胞から再びせり出して眼胞を覆った部分の中央が外側に向かって膨らみ水晶体前半となる。頭足類ではこれら2つが融合して水晶体となる。ただし他の大多数の無脊椎動物の眼と同様に「背向性眼」(=反転眼)ではない。発生においては無脊椎動物の眼はすべてが表皮由来であるが、脊椎動物の外側眼は網膜組織と色素上皮層の2層構造などが脳由来であり角膜の外層と水晶体が表皮由来である。脊椎動物の頭頂眼は脳由来の網膜だけの1層構造である。また脊椎動物の外側眼は背向性眼(反転眼)であり、視神経乳頭による盲点があるが、頭頂眼は反転眼ではない[6]。
目の形態
眼を形態で大きく分類すると単眼・複眼・カメラ眼の3つに分けられる[8]。
単眼
光受容細胞が杯状の構造を形成し、その外層にレンズを備える構造を単眼と呼ぶ。単眼は、レンズと網膜を備えるが、ピント調節や絞りなどの機能はない。環形動物、多くの軟体動物、節足動物は単眼を持つ。
複眼

トンボにみられる複眼
複眼は、個眼の集合体である[8]。この個眼がたくさん集まり半球状に配列されたものが複眼である[8]。
個眼は外層から個別のレンズ、円錐晶体、視細胞層から成り立つ。個眼同士は光を通さない隔壁で分かれている。視細胞層はミツバチの場合、外部からの光を直接受ける中心の感棹と周囲に8つ並ぶ光受容細胞からなる。光受容細胞はミツバチの場合、紫外線に最も高い感度をもつもの2つ、青に感度を示すもの2つ、緑に感度を示すもの4つから成り立つ。
レンズを小さくすることで焦点距離を短く出来るため、体のスペースがほとんどない小さな生物に適した構造である。また動きを捉えることに適している[8]。その一方で、対象を精細に見る上では限界がある。画像を捉えるための細胞が、人間では網膜上に1億以上あり脳への伝達細胞も約100万ある。しかし複眼は各個眼をそれほど多く備えることが難しく、比較的多いトンボでも数万でしかない[8]。また遠くのものを見る機能にも劣り、昆虫の視力は0.01程度でしかない[8]。
カメラ眼
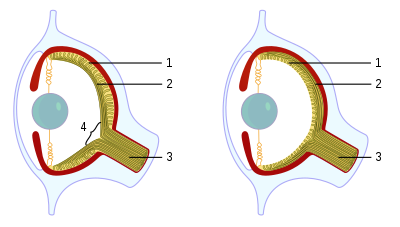
脊椎動物(左)とタコ(右)の眼: 脊椎動物の網膜では、視細胞(1)は網膜の内側に、神経節細胞(2)は網膜の外側にある。神経節細胞の軸索は視神経(3)となるが、視神経が網膜から出る部位には視細胞は存在せず盲点(4)となる。タコの眼では視細胞は網膜の外側にあるため、盲点は存在しない。
カメラ眼とは、ピントの調整を行える可動な1枚の大きなレンズまたは水晶体、取り込む光量を調整する機能、そして網膜のような面積が広い光受容組織を供えた、いわゆるカメラと基本的に同じ構造を持つ目である。これは大型の動物でも比較的小さな目で視力を持つのに適し、すべての脊椎動物が備えている[8]。角膜やレンズを持った事で像を結ぶ点に優れ、さらにレンズを動かす筋肉を発達させたため遠くを見る事でも有利である[8]。
また、軟体動物門頭足綱のタコやイカは、いわゆるカメラ眼を持つ[8]。カメラ眼はピント調節が可能である点では、脊椎動物の眼光学系に類似する。しかし、網膜の構造や機能は脊椎動物とは大きく異なる。脊椎動物の網膜は脳や神経になる体内の層が進化して形作られたが、タコなどの光受容体は皮膚層がくびれて体内に取り込まれて形成された[8]。たとえば、脊椎動物の視細胞は光に過分極応答するが、イカやタコの視細胞は光に脱分極応答する。
水棲と陸棲
魚類の眼は水圧と低屈折率という点で水中での生活に適した構造を備えている。水圧が眼の組織を強く押してくるため強固になっている。ある種の魚類は眼球の強膜に軟骨や骨を持っている。また、水は角膜や水晶体の屈折率に近いため陸上生物のような眼のレンズでは光線があまり屈折せずに網膜へ届いてうまく像を結べなくなる。魚眼レンズという言葉があるように、魚の眼は水中での低い屈折率を補うために水晶体はレンズ状では済まなくなり球形をしている。ヒトを含むほとんどの陸棲動物の眼は遠近のピントを合わせるのに水晶体の厚みを変えているが、魚の眼は球状の水晶体でありカメラのレンズがピントを合わすように、眼球の中で前後に移動している。遠くの物を見る時には「水晶体牽引筋」を収縮させて水晶体を眼の奥の網膜方向へと引き、近くの物を見る時には弛緩させて水晶体を眼の前方へ戻す。哺乳類のイルカやアザラシ、そして鳥類でも水鳥は陸上でも水中でも良好な視力が求められるが、かれらは柔軟な水晶体を毛様体筋で強く引くことで陸上ではレンズ状/水中では球状になるよう大きく変形させている。鳥類でもペンギンの眼は水中にのみ対応しており、陸上では極度の近視になってあまり周囲が見えていない。
陸棲動物では角膜を乾燥や汚れから守るために瞼が発達したが、水棲動物は瞼が無いかあっても簡素なものである。両生類は上下に瞼を持ち、さらに薄く透明な瞬膜(しゅんまく)がある。瞬膜は鳥類も備えており、飛行中は瞼を開いたまま瞬膜を閉じて乾燥から守られ外を見ることができる。陸棲動物は角膜を潤すために眼窩腺を持つ。両生類、爬虫類、鳥類ではハーダー腺が発達している。哺乳類では眼窩腺の1つに涙腺がある[6]。
眼の配置

眼の進化
眼の型、構造は様々であるが、その配置はある程度の共通性がある。ある程度以上の視力を持ち、それに多く頼って生活していると見られる動物では、主要な眼を一対、ある程度同一方向へ向けているのが普通である。例えば同じカメラ眼でも起源の異なる脊椎動物と頭足類、複眼を持つ昆虫や大型甲殻類はすべてこのような配置の眼を持つ。両眼で一方向を見ることで立体視が可能になり、両眼視差を利用することで距離情報がより正確になる。例を挙げれば草食動物のウマやウサギは目の間隔が離れており視野角が広く、迫り来る敵を察知しやすくなっている。その反面ライオン等の肉食動物は目の間隔が狭く、前方の獲物の距離を正確に認識することができる。
クモ類では8個の眼が並んでいるが、ハエトリグモのように視覚に頼った狩りをするものでは2眼が特に良く発達する例が見られる。小型の動物では目の位置を出来るだけ高くするものもあり、クモ類ではハエトリグモ類やササグモ科など、徘徊性に特に適応したものでは頭部が特に盛り上がり、カニ類では眼だけが体から上に伸ばされている。
水中と陸上では眼の働き方も異なる。ワニやカバ、カエルのように陸上生活を行いながら水にたびたび入る動物は、目は鼻とともに頭の上部に位置する共通の適応が見られる。また、ヨツメウオやミズスマシのように水面にあって水上と水中を同時に見る動物では、上下の世界をそれぞれ別の目で見る例もある。
ツバメなどの鳥類は中心窩が2つあり、両目で見た場合の前方1箇所の他に、左右の片眼だけそれぞれ側方の1箇所が良く見える点がある。円口類のヌタウナギは光の届かない300m程の海中に適応したため、祖先が持っていた眼は直径1mmほどまで小さくなり水晶体や虹彩は退化して失われて皮下へ移動してしまい、明暗を判別できるだけになっている。同様に、洞窟、深海、土中のような光の無い世界に適応した動物では、元からあった眼が退化してしまったものが多く存在する[6]。
視覚
視力
目がものを捉える力を視力という。これは具体的には、空間にある2点を識別する能力であり、眼球の屈折や調整能力の低下、または眼球の透過性や網膜の異常、そして視覚伝達神経の不備などがあると低下する[9]。
色覚
光を捉える目の細胞には、明るい場所で働く錐体細胞と、暗い場所で使われる桿体細胞がある。色を認識するのはこのうち錐体細胞であるが、全ての色すなわち全ての波長の光を捉えられない。人間の目が識別可能な波長幅を特に可視光線と言うが、見えている光の波長は生物によって異なる[8]。
全ての目において光を捉える事は、視細胞に蓄えられたオプシンというタンパク質分子が最初に光を吸収する事で始まる。このオプシンには複数の種類があり、それぞれ受け止める波長が短い方から「紫外/青型」「青型」「緑型」「赤/緑型」の4種に分けられる。この4種は、生物の中で5億年前頃に揃ったと言われる[8]。このうち人類が持つオプシンは、波長が青寄りになり420nm付近に特化した「紫外/青型」と、530nm付近に特化した「赤/緑型」(緑)、560nm付近に特化した「赤/緑型」(赤)の2種類3サブタイプであり、色覚は3色型となる[8]。
脊椎動物のオプシンを調べると、魚類・爬虫類・鳥類が4種類を持っており、両生類は見つかっていないがこの例に当たると考えられている。特に魚類はサブタイプも多くメダカは8タイプを持つ。ところが進化上で、ほとんどの哺乳類は「青型」「緑型」を失った。これは初期の哺乳類が主に夜行性だった事が影響したと考えられる。さらに「紫外/青型」のうち紫外線を見る能力も失うが、例外的に有袋類のオポッサムや齧歯類の中には紫外線を感知する目を持つ者もいる[8]。
オプシンを失い2種類2サブタイプの色覚(2色型)となった哺乳類のうちから、霊長類は進化の過程で「赤/緑型」のサブタイプを増やした。これは、主な霊長類が生きた場所が森林であり、生存する上でものを見分ける際に色が重要だったためと考えられる[8]。中には特殊な例もあり、オマキザルはサブタイプが異なる2色型と3色型が群れの中で混在するケースが見つかっている[8]。
眼の進化
生物がいつ視覚的な能力を獲得したのかは定かでなく、一説には21億年以上前の単細胞生物が光を感知できたというが定かではない[8]。これは柔らかい構造である目は化石として残りにくいためである。それでも先カンブリア時代以前から、生物は光を捉える表面細胞を備えていたと考えられる。これが爆発的な進化を遂げたのがカンブリア紀(約5億2000万年前)のいわゆるカンブリア爆発である。浅瀬の海を舞台に、視力を持てば捕食のためまたは敵から逃れる上で非常に有利に働く。カンブリア紀は、目が生存に有利な機関として進化と多様化を進めたと考えられ、これを「光スイッチ説」と言う[8]。
チャールズ・ダーウィンは、複雑な目の構造が自然にできあがるという考えは合理的でないように思われると述べたが、それでも自説を放棄せず、目も生物は進化の中で獲得したという説を堅持した[8]。近年のシミュレーションでは、生物の世代交代を1年とした場合、単純な目がカメラ眼に進化するまでに必要な時間は40万年以下という結果も得られた[8]。
進化論にまつわる議論の歴史として、「脊椎動物の複雑な眼の構造の、どれか1つでも要素が欠けると正常な視力が得られないと考えられるとし、また「最初から完全な状態で作られていなければ眼は眼たりえない」、「すると自然選択で有利とならないので、目が発生したことが説明困難」とするような説が、繰り返し指摘され、進化論をめぐる難問のひとつとみなされてきた歴史がある。上記のように最近では「現実には各種動物において様々な型の眼が見られ、それらの性能もまた多様である。高度なものではヒトと同様かそれ以上の情報を提供するとされるものもあり、逆に簡単な明暗程度しかわからないであろうものもある」ともされ、“完全な眼”を想定するのは困難であり、また、不完全な視力では役に立たないとの論旨も根拠を持たないと言える。
脚注
- ^ abcdefghijk解剖学第2版、p.148、第9章 感覚器系 1.視覚器
^ 解剖学第2版、p.135-146、第8章 神経系 4.末端神経系
- ^ abcdef佐藤・佐伯(2009)、p.245-247、第12章 感覚、2.視覚visual sensation、1)眼球の構造と働き
^ 医学大辞典(医学書院、ISBN 4-260-13651-8)
^ 目の愛護デー(日本眼科医会)
- ^ abcdefgh岩堀修明著、『感覚器の進化』、講談社、2011年1月20日第1刷発行、ISBN 9784062577
^ 頭足類では脊椎動物の眼のように瞳孔、角膜、瞼まで備える。
- ^ abcdefghijklmnopqrstニュートン (2012-6) p.94-99 知られざる眼のヒストリー
^ 佐藤・佐伯(2009)、p.249-251、第12章 感覚、2.視覚visual sensation、3)視覚の性質と調整
参考文献
- 編集長:竹内均「ニュートン2012年6月号、雑誌07047-06」、ニュートンプレス、2010年。
- 河野邦雄、伊藤隆造、坂本裕和、前島徹、樋口桂 『解剖学第2版』 財団法人 東洋療法学校協会、医歯薬出版、2006年、第2版第1刷。.mw-parser-output cite.citation{font-style:inherit}.mw-parser-output .citation q{quotes:"""""""'""'"}.mw-parser-output .citation .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .citation .cs1-lock-limited a,.mw-parser-output .citation .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .citation .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-ws-icon a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/4/4c/Wikisource-logo.svg/12px-Wikisource-logo.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-maint{display:none;color:#33aa33;margin-left:0.3em}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
ISBN 4-263-24207-6。 - 監修:佐藤昭夫、佐伯由香 『人体の構造と機能 第2版』 医歯薬出版、2009年、第2版第6刷。
ISBN 978-4-263-23434-1。
関連項目
- 視覚
- 視力
- まぶた
- 眼科学
- 眼鏡
外部リンク
視覚 ‐目の構造 (ビジュアル生理学 内の項目)


