種子
この記事は検証可能な参考文献や出典が全く示されていないか、不十分です。 出典を追加して記事の信頼性向上にご協力ください。(2011年5月) |

オオトウワタの種子
種子 (しゅし、〈たね〉) とは種子植物で有性生殖によって形成される散布体である。一般には、単に種(たね)と呼ばれることが多い。
種子は親植物の組織起源の種皮(しゅひ)という皮に包まれ、その中には受精卵から発育した幼い植物体、すなわち胚が入っている。種子はめしべにある胚珠から発達する。花のつくりの場合は、子房は柱頭の下などに多い。被子植物の場合、種子は子房に包まれていて、これが後に果実となる。裸子植物の場合は、めしべの表面に乗っている。
農業上繁殖に用いられるものは、厳密には種子でなくとも、種もみ・種芋・種馬などと呼ばれる。
目次
1 種子の構造
2 種子の散布
3 種子の休眠と発芽
4 種子の起源
5 関連図書
6 脚注
6.1 注釈
6.2 出典
7 関連項目
8 外部リンク
種子の構造
たとえば、豆はさまざまなマメ科植物の種子である。これを例に説明する。
- 豆は、普通、細長い鞘の中にいくつか並んで収まっている(アメリカデイゴの豆果)。この鞘は、めしべの基部、子房が膨らんだものである。子房は胚珠を中に収め、胚珠が種子に発達してもそれを包んでいる。子房に由来する構造を果実という。
- 個々の豆の表面は、薄い皮に包まれている。この皮を種皮(しゅひ)と呼ぶ。種皮は珠皮に由来する。
- 種皮をはがすと、中には大きく2つに割れる部分がある。半球形の2つの部分は、小さな出っ張りの部分でつながり、その間には小さな折り重なったような構造がみられる。半球形の部分は子葉(しよう)と呼ばれ、発芽すると双葉(ふたば)として地上に姿を現す(種子内に留まるものもある)。丸く膨らんでいるのは、栄養が蓄えられているからである。
- 子葉をつなぐ出っ張りは、胚軸(はいじく)と呼ばれ、芽生えた苗の茎になる部分である。胚軸に続く部分(子葉と反対側)が根となる幼根である。間に挟まった折り重なったものは、双葉の次の本葉が既に形成されているのが見えているものである。
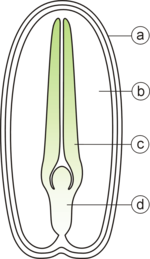
双子葉植物の種子の断面模式図。a:種皮 - b:胚乳 - c:子葉 - d:胚軸
このように、種子の中には、既にこれから発芽する苗が含まれている。
カキの種子では、中の様子がかなり異なっている。種子を半分に切ると、幼い植物体は種子に比べてずっと小さく、種皮の中には半透明の固いものが詰まっている。これは胚乳と呼ばれ、胚嚢(はいのう)の中央細胞(2つの極核を含む)と花粉管内に生じた精細胞の1つが受精したもの(核としては2個の極核と1個の精核、合計3つの核が受精する)に由来する。このような種子では、胚乳に栄養が蓄えられている。イネやムギでは幼植物は種子の末端部分にあり、種子本体の大部分を胚乳が占める。
基本的に被子植物の種子は、カキの例のように、発生の出発点において極核を含む中央細胞と精細胞が受精した3倍体核に由来する胚乳を生じるが、マメ科など一部の植物では二次的に胚乳が退化し、かわりに子葉などに発芽のための栄養分を蓄える。ほかに、ラン科(エビネの種子)など若干の植物で、発芽のための蓄えを持たず、菌類との共生に依存するなどして発芽する例がある。裸子植物の胚乳は1倍体の雌性配偶体自体に由来し、シダ植物の前葉体と相同の器官で、被子植物の胚乳とは異質な面がある。
多くの種子は、幼植物が発芽するための栄養分を子葉か胚乳に蓄えているため、動物から見れば魅力的な食料である。人間の食物の中にも種子はさまざまに用いられ、特にマメ科やイネ科のものは、しばしば主食の位置を占める。主食に使われるイネ科の種子を穀物と呼ぶ。蓄えられる栄養分は、デンプンか油脂の形である場合が多い。
種子の散布
植物には基本的に移動能力がない。ある場所で種子が発芽をすれば、そこに一生とどまるのが基本である。従って、種子が好適な場所に到達する何らかの手段がなければならない。現に親植物が生息している以上、親の根元は好適地であるから、親が自分の根元に種子を落とすのは一つの方法だが、それでは親子あるいは子供同士で無意味な競争をせねばならない。また分布拡大の意味からも、種子はある程度以上、遠くに運ばれる必要がある。
実際には、種子にも移動能力はないので、種子の散布en:Seed dispersalは何か外の力に頼らざるを得ない。そのためそれぞれの植物は、何かに頼って種子を散布するための方法を発達させてきた。

サトウカエデの種子
- 風による散布
- 物理的な力に頼るものとしては、よく見られるものの一つである。裸子植物のクロマツやアカマツでは、種子の一端が薄い膜状に伸びており、空中にでると風を受けて、回転しながら飛んでゆく。同じような構造を発達させたものに、カエデ科のもの(モミジの仲間)やアオギリなどの果実がある。同じ風を利用するにも、キク科のタンポポ(セイヨウタンポポの痩果)などは、果実の一端から多数の毛を生じて、これが風を捉える方法を取っている。同様なものは、ガガイモ科のガガイモの種子、イネ科のススキの果実など、多くのものに見られる。
- 水による散布
- 水はものを運ぶ力が強く、特別な適応がなくても勝手に運んでくれるので、多くの種がその恩恵をこうむっていると思われる。特に、水による運搬への適応を示しているので有名なものにラッカセイ(落花生・ピーナッツ)がある。豆の鞘が空気を含み、水に運ばれやすくなっている。スゲの仲間の果実は後述のアリによる種子散布に適応したものと水による種子散布に適応したものの2つに大別される。
- 海流による散布
- 海岸性のごく限られた植物に見られる。陸上植物には海水が有害なので、まず塩分に耐えられる事が前提になる。ハマユウやマングローブの胎生種子や、ゴバンノアシなど、熱帯の海岸性植物には、大きく膨らみ海水に浮かぶ果実や種子をつけるものがあり、これらは海流による散布に適応したものである。ココヤシの果実は遠い海岸に流れつくことで有名である。
- 動物による散布
- これにはいくつかの型がある。
- 餌となることによる散布を目指すものは、種子や果実が動物の食料として選ばれ、この時に散布の手助けをしてもらうことを期待する。
- いわゆる果物(くだもの)を多く含む漿果の果肉はそのために発達したものである。哺乳類や鳥類などに果物を食わせておき、同時に種子を丸呑みさせ、糞と一緒に排出され、そこで発芽する。この場合、果肉は大きく柔らかく、糖分や脂肪を多く含む。それに対して種子は小さかったり、大きくて硬く、壊されにくい構造になる。ドリアンやレイシ、イチイのように果皮起源の果肉ではなく仮種皮を果肉として発達させるものも多い。
ドングリの場合、食料になるのはデンプンを多量に蓄積した種子そのもの(コナラの堅果)である。種子散布に寄与する動物はリスのように種子を集めて貯蔵する習性がある動物である。餌になるとその時点で種子としての役割を失うが、それでも絶滅することがないのは、壊されるのが子葉の一部に過ぎなければ充分発芽に役に立つという側面があるとともに、貯蔵種子の一部を忘れてしまったり、食べ残したりしたものも種子散布に役立っていると考えられている。- 種子そのものが食料になるのではなく、種子に餌をつけて運ばせるように進化したものがある。スミレ類やカタクリは種子に付属するエライオソームと呼ばれる脂質を多く含む肉質の部分を持つ(ニシキスミレの種子)。これがアリの餌となってアリに運ばれる。
- 動物の体表面にくっつき、運んで貰うための種子を発達させたものもある。果実や種子の一部に粘着物質を出したり、棘や毛で絡み付いたりするようになっているものである。人間の衣服にもよくくっつき、結実期(日本では主に秋)の山野にでかければ、必ず何種類かの種子に絡み付かれ、後で取るのに苦労する、いわゆる「ひっつき虫」イガオナモミの「いが」がこれにあたる。
- 餌となることによる散布を目指すものは、種子や果実が動物の食料として選ばれ、この時に散布の手助けをしてもらうことを期待する。
- 機械的に種子を飛ばす仕組みを発達させたもの
- 有名なのはホウセンカで、成熟した果実は何かの刺激があると割れて、皮が大きくゆがみ、中の種子を跳ね飛ばすようになっている。

服に引っ付いたタウコギの種子

はじけたツリフネソウの仲間の果実
 メディアを再生する
メディアを再生する
バクダンウリの果実



種子の休眠と発芽
種子は、内的な要因で発芽が阻害されている状態(休眠状態)では好適な条件下に置いても発芽しないが、休眠が解除された状態では、好適な条件下に置かれると、発芽過程が始まる。呼吸によるエネルギー生産が始まり、種子の中にある幼い植物体が成長をはじめ、種皮を破って伸び、葉を地上に現す。これを発芽と言う。
多くの植物では、種子の中の植物体は休眠状態にある。ただし、休眠に入る機構、休眠が維持される機構、休眠が打破される機構は複雑であり、解明されていない部分も多い。一般に、発芽過程を開始していない(生命活動を停止している)種子は、長い期間にわたって生き延びられることが知られている。ただし、生き延びられる期間は種によって様々であり、何年も保たないものもあれば、数十年にもわたって発芽力を維持するものもある。長生きで有名なのはハスで、日本では弥生時代の遺跡から発掘された種子が発芽した例があり、「大賀ハス」として知られている*。
(*)長い間地層(日本考古学では「土層」)中に保存されていた数千年前のハスが発芽し、いわゆる古代ハスと呼ばれているものは、ハスの種子そのものを年代測定した例はない。これらは同じ層位や上下の層位の他の物質(土壌有機物や木片など)を測定したものから推定したものである。発芽した種子は現生種子の混入の可能性を排除できず、根拠としての確実さは乏しい。種子を直接測定した例では、今のところ500年程度の寿命があることが分かっている。
種子は、好適な条件下で発芽するが、野生種の場合、種子の集団が一斉に発芽するわけではなく、発芽せずに残るものがあるという。これは、発芽の条件が整っても、成長の過程で条件が悪化すれば枯れるので、全種子が死滅する危険を回避するための適応であると言われる。栽培植物の場合、発芽や収穫の効率を上げるため、休眠性を失い、好適な条件下で一斉に発芽するように品種改良されている。
どの様な条件で、休眠が解除され発芽が始まるかは、その種の性質によりさまざまである。
種子の起源

シダ植物の胞子のう
種子を形成した最初の植物は古生代末期のシダ種子植物とされている。 化石として葉身表面に種子を並べたようなシダ植物的なものが発見されており、この仲間では杯状の種皮の中に、雌性前葉体が閉じこめられている。
普通のシダ植物は、葉の上に胞子を形成し、放出された胞子は湿った地面で発芽して前葉体となる。前葉体の下面に造精器、造卵器を形成、受精が起こる。受精卵は前葉体の上で発生を始め、植物体が成長する。
進化の道筋としては、恐らくそのような形から、精子のみを作る雄性前葉体と卵のみを作る雌性前葉体が分かれるものが現れたと考えられる。現在でも水生シダ類などにそのようなものがあり、雌性配偶子になる大胞子と雄性配偶子になる小胞子を別々の胞子のうの中に作る。
更に、大胞子が胞子のうから出る前に発生を始めるものが現れ、それを保護するための覆いが発達したのが種子の起源であると考えられる。このようなことになったのは、陸上生活する維管束植物の生活史の中で、前葉体の時代が最も水に依存するからと推定される。精子が卵の所まで泳がなければならない。そのためには水が必要になり、水がなければこの段階を超えられない。そこで、この段階を母植物の上で過ごしてしまう方向へ進化が進んだのであろう。精子を作る小胞子は、種子のそばで発芽しそこで精子を作れば、母植物の葉先の水滴だけで受精が可能になる。

ソテツの雌花
種子の皮である種皮は、胞子のうの袋とそれを守る皮からできている。未発達の種子を胚珠と呼ぶ。
受精した卵は母植物から栄養をもらってその場で発生をはじめ、小さな植物体にまで発達して休眠状態となる。そして種子が放出され、好適な場所に落ちるまでを待つことになる。
現生の裸子植物では、大胞子のうをつける胞子葉はごく簡単な形のものが多く、ソテツの雌花に少しだけ葉の形の面影が見られる。それ以外のものでは鱗片状の形で、それが軸の周りに密に折り重なり、いわゆる松ぼっくりの形になっている。鱗片1枚を取り出すと、その上に左右1つずつの胚珠が並んでいる。
さらに、胚珠を胞子葉が包むようになったのが被子植物である。胚珠は子房という袋状の部分におさまり、小胞子(花粉)は胞子葉(めしべ)の特定の部分(柱頭)に付着して、胚珠まで花粉管を伸ばすことになった。
関連図書
- 「タネの大図鑑」(監修:サカタのタネ 発行:PHP研究所)
脚注
注釈
出典
関連項目
- 種子植物
- 主要農作物種子法
散布体 - 胞子
- 果実
播種 - 発芽 - 芽
- 花粉
外部リンク
樹のタネ図鑑 - 樹の種の写真集 (群馬県山林種苗緑化協同組合)
日本の帰化植物 種子画像データベース (岡山大学 資源生物科学研究所 野生植物研究室) ※インターネットエクスプローラのみ閲覧可能
日本の帰化植物 種子画像データベース 上記ページの Internet Explorer 以外のブラウザ対応ページ。検索など一部機能が使えないとのこと
| |||||||||||||||||||||



