動物
| 動物界 | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
生息年代: エディアカラ紀 - 現世 PreЄ Є O S D C P T J K Pg N | ||||||||||||||||||
 各画像説明[注釈 1] | ||||||||||||||||||
分類 | ||||||||||||||||||
| ||||||||||||||||||
| 門 | ||||||||||||||||||
本文参照 |
動物(どうぶつ、羅: Animalia、単数: Animal)とは、
生物学において生物の系統群の一つ。「動物」という言葉がつく分類群名としては後生動物や原生動物があるが、本稿でいう「動物」は後生動物の方を指す。後者は進化的に異なる雑多な生物をまとめたグループ(多系統群)であり、いずれも後生動物とは別系統である。- 日常語において、動物とは1.の意味の動物のうち、ヒト以外のもの[1]。特に哺乳類に属する生物を指す事が多い[1]。
本項では1の意味を解説する。
目次
1 概要
1.1 特徴
2 動物の細胞
2.1 細胞小器官
2.2 細胞外マトリックス
3 生殖、発生、分化
3.1 生殖
3.2 発生
3.3 細胞分化と器官形成
4 起源と進化
4.1 起源
4.2 古生物
4.2.1 先カンブリア時代
4.2.2 古生代
4.2.3 中生代
4.2.4 新生代
4.3 化石動物についての動物門
4.4 絶滅した動物
5 現生の動物の系統
5.1 系統
5.2 前左右相称動物
5.3 左右相称動物
5.3.1 冠輪動物
5.3.2 脱皮動物
5.3.3 新口動物
6 参考文献
7 脚注
7.1 注釈
7.2 出典
8 関連項目
概要
動物は、哺乳類、爬虫類、鳥類、両生類、魚類といった脊椎動物はもちろん、貝類、昆虫、サナダムシ、カイメンなど、幅広い種類の生物を含んだ系統群である。
20世紀末の分子遺伝学の知見を踏まえると、生物は真正細菌、古細菌、真核生物の3つに分かれるが(3ドメイン説)[2][3][4]、動物はそのうちの真核生物に属し、他に真核生物に属するものとしては植物、菌類(キノコやカビ)、原生生物が挙げられる。なお、原生生物の一部である原生動物(ゾウリムシ、ミドリムシ、アメーバ等)は本稿で言う動物(後生動物)とは別系統であり、しかも多系統である事が判明している。
なお、初等教育では3ドメイン説以前の二界説(2011年まで)ないし五界説(2012年以降)に基づいて生物の分類を説明している[5]。二界説に基づいた説明では原生動物を「動物」とみなしていたが、すでに述べたように3ドメイン説では(後生)動物とは別系統であるとみなされている。一方、五界説での動物は3ドメイン説のものと基本的に同じであり、原生動物は原生生物として動物とは区別されている。
動物は真核生物の中ではオピストコンタ(後方鞭毛生物)という単系統性が強く支持されている系統群に属し、ここには動物以外に菌類や一部の真核生物が属する。オピストコンタに属する生物は、後ろ側にある1本の鞭毛で進むという共有形質を持ち、動物の精子やツボカビの胞子が持つ鞭毛がこれにあたる。
さらにオピストコンタにはホロゾアという系統群と、Holomycotaという系統群があり、動物は前者、菌類は後者に属する。なお動物の起源とされる(後述)襟鞭毛虫もホロゾアに属する。
特徴
多細胞の真核生物である[6][7]
従属栄養生物である[7][8]。すなわち植物のような独立栄養生物と違い、無機物から自力で栄養源を得る事はできない。- 非常に少数の例外的な動物を除き、好気呼吸する[9]。すなわち酸素を使った細胞呼吸をする。
運動性がある[10]すなわち、自発的に体を動かす事ができる。ただし生涯の途中で付着生物と化すなど、運動性がない時期がある動物もいる。- ほとんどの動物には、胚発生の初期に胞胚という段階がある[11]。
動物の細胞
動物の細胞は、全ての真核生物の細胞に共通した以下の構造を持つ。
細胞膜:細胞を包んでいる膜[12]。内部は生体物質を含む水溶液があり代謝の場となっている。リボソーム、細胞質(原形質)といった共通の構成要素を持っている。
DNA:塩基配列または遺伝暗号 (genetic code)と言うヌクレオチドの塩基部分が並ぶ構造を持ち[13]、遺伝情報の継承と発現を担う。真核細胞のDNA は、一本または複数本の分子から構成される直線状で原核生物よりも多く[14]、染色体と呼ばれる[15]。
細胞質:細胞の細胞膜で囲まれた部分である原形質のうち、細胞核以外の領域のこと。真核細胞の細胞質には細胞骨格(サイトスケルトン)と呼ばれる微小な管やフィラメント状がつくる網目もしくは束状をした3次元構造[16]がある。これが特に発達した動物の細胞では、細胞骨格が各細胞の形を決定づける。
細胞小器官
典型的な動物細胞には、以下のような細胞小器官がある(番号は図のものと対応):

典型的な動物細胞の模式図
核小体(仁):細胞核の中に存在する、分子密度の高い領域で、rRNAの転写やリボソームの構築が行われる。
細胞核:細胞の遺伝情報の保存と伝達を行う。
リボソーム:mRNAの遺伝情報を読み取ってタンパク質へと変換する機構である翻訳が行われる。
小胞:細胞内にある膜に包まれた袋状の構造で、細胞中に物質を貯蔵したり、細胞内外に物質を輸送するために用いられる。代表的なものに、液胞やリソソームがある。
粗面小胞体:リボソームが付着している小胞体の総称。
ゴルジ体:へん平な袋状の膜構造が重なっており、細胞外へ分泌されるタンパク質の糖鎖修飾や、リボソームを構成するタンパク質のプロセシングに機能する。
微小管:細胞中に見いだされる直径約 25 nm の管状の構造であり、主にチューブリンと呼ばれるタンパク質からなる。細胞骨格の一種。細胞分裂の際に形成される分裂装置(星状体・紡錘体・染色体をまとめてこう呼ぶ)の主体。
滑面小胞体:リボソームが付着していない小胞体の総称。通常細管上の網目構造をとる。粗面小胞体とゴルジ複合体シス網との移行領域、粗面小胞体との連続部位に存在する。トリグリセリド、コレステロール、ステロイドホルモンなど脂質成分の合成やCa2+の貯蔵などを行う。
ミトコンドリア:二重の生体膜からなり、独自のDNA(ミトコンドリアDNA=mtDNA)を持ち、分裂、増殖する。mtDNAはATP合成以外の生命現象にも関与する。酸素呼吸(好気呼吸)の場として知られている。また、細胞のアポトーシスにおいても重要な役割を担っている。mtDNAとその遺伝子産物は一部が細胞表面にも局在し突然変異は自然免疫系が特異的に排除[17] する。ミトコンドリアは好気性細菌でリケッチアに近いαプロテオバクテリアが真核細胞に共生することによって獲得されたと考えられている[18]。
液胞:電子顕微鏡で観察したときのみ、動物細胞内にもみられる。主な役割として、ブドウ糖のような代謝産物の貯蔵、無機塩類のようなイオンを用いた浸透圧の調節・リゾチームを初めとした分解酵素が入っており不用物の細胞内消化、不用物の貯蔵がある。
細胞質基質:細胞質から細胞内小器官を除いた部分のこと。真核生物では細胞質基質はどちらかと言えば細胞の基礎的な代謝機能の場となっている。
リソソーム:生体膜につつまれた構造体で細胞内消化の場。
中心体:細胞分裂の際、中心的な役割を果たす。
細胞外マトリックス
動物の細胞はコラーゲンと伸縮性のある糖タンパク質からなる特徴的な細胞外マトリックスで囲まれている[19]。細胞外マトリックスは細胞外の空間を充填する物質であると同時に、骨格的役割(石灰化による骨、貝殻、海綿骨針といった組織の形成[20] )、細胞接着における足場の役割(例:基底膜やフィブロネクチン)、細胞増殖因子などの保持・提供する役割(例:ヘパラン硫酸に結合する細胞増殖因子FGF)などを担う。また動物細胞は、密着結合、ギャップ結合、接着斑などにより細胞結合・細胞接着している[21]。
海綿動物や平板動物のような少数の例外を除き、動物の体は組織に分化しており[22]、組織としては例えば筋肉や神経がある。
生殖、発生、分化
生殖

トンボの交尾
ほぼ全ての動物は何らかの形で有性生殖を行い[23]、その際減数分裂により一倍体の配偶子を作る。2つの配偶子が融合する事で新しい個体が生まれるが、この場合小さくて運動性がある配偶子を精子、大きくて運動性を持たない配偶子を卵子といい[24]、配偶子が融合する過程を受精、受精の結果できあがった細胞を受精卵という[25]。また精子を作る個体をオス、卵子を作る個体をメスという。一つの個体が精子と卵子を両方作れる場合は雌雄同体であるという。
発生
一倍体である精子と卵子が受精する事で、二倍体の受精卵が形成される。この際精子由来のミトコンドリアは酵素により分解されるので[26]、ミトコンドリアは卵子からしか受精卵に伝わらない。
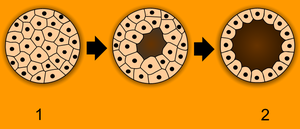
胞胚形成: 1 桑実胚、2 - 胞胚
受精卵は卵割という細胞増殖を繰り返す事で多細胞の胚を形成する[26]。一般的に卵割の際は核は複製されるが細胞質は卵細胞のものを分割して使うという特徴がある[26]。一般的に、卵割が進むと胚の内部に隙間が形成され、この隙間が広くなったものを胞胚腔といい、大きな胞胚腔が形成される時期の胚は胞胚と呼ばれる[27]。なお、昆虫や両生類など多くの動物では、卵割期の細胞増殖を急激に行うために通常の細胞分裂で行われる一部の過程(G1期とG2期の過程)が省略され[28]、胞胚中期になるとこの省略が終わる(中期胞胚転移)[29]。それに対し胎生の哺乳類ではこういった省略は起こらない[29]。
胚が形成される過程で、体軸という体の向きが決定がなされ、その向きには前後軸、背腹軸、左右軸などがある[30]。例えば両生類では精子が受精した位置により背腹軸が決まり、受精した側が腹側、その反対側が背側になる。
一部の原始的な動物(前左右相称動物[注釈 2])以外は胞胚後期になると胚葉が形成される(形態形成運動)[31][32]。胚葉には将来消化管になる内胚葉、将来表皮や神経系などになる外胚葉、そして体のそれ以外の所(例えば体腔、循環系、内骨格、筋肉、真皮)になる中胚葉の3種類がある。典型的にはこの際内胚葉の部分が陥没し、原腸が構成される。この時期の胚を原腸胚という[注釈 3]。
なお、前左右相称動物の場合は、海綿動物のように胚葉が形成されないものや[33](無胚葉性の動物と呼ばれる)、内胚葉と外胚葉の2つのみしか形成されない動物(二胚葉性の動物)もいる[注釈 4]。これに対し胚葉が3つとも形成されるものを三胚葉性の動物という。
脊椎動物などではこの後神経管が形成される神経胚期へと進む。例えばニワトリでは、外胚葉に神経板という領域ができ、それが胚の内側に丸まる事で神経管ができ、さらに直下に脊索が形成される[34]。神経管の前方には前脳、中脳、後脳という3つの膨らみが形成され、これらが将来脳になる[35]。
細胞分化と器官形成
脊椎動物などでは、組織や器官を形成するため、胚細胞が特定の機能を持った細胞に変化する(細胞分化)[36]。この際、基本的な細胞機能の維持に必要な遺伝子(ハウスキーピング遺伝子)の機能は残しつつ、特定の機能に必要な遺伝子を新たに発現し、逆に分化後には不必要になる遺伝子をDNAメチル化により不活性化する[36]。
脊索の両側の沿軸中胚葉から体節が形成され、体節と隣接した外側の中間中胚葉からは腎節が形成される[37]。
体節はやがて皮節、筋節、硬節に分かれ、これらはそれぞれ皮膚の真皮層、骨格筋、椎骨などが形成され[37]、腎節からは腎臓や生殖腺が形成される[37]。
中間中胚葉のさらに外側には予定心臓中胚葉という、将来心臓関連の組織になる部分があり、これは壁側中胚葉と臓側中胚葉に転移する[38]。前者からは体腔を覆う胸膜や腹膜が形成され、後者からは心筋、平滑筋、血管、血球などが形成される[39]。
心臓は生命の維持に不可欠なので、発生の早い段階で中胚葉から形成される[40]。なお、予定心臓中胚葉は中胚葉の正中線を隔てた両側に2つ存在するが、これら2つは移動して胚の前方で合流して心臓を形成する[40]。
脊椎動物では外胚葉と中胚葉の相互作用で四肢が形成される[41]。ヒトの手足は水鳥と違い、指の間に水かきがないが、これはアポトーシスの作用で水かき部分の細胞を「自殺」させている為である[42]。
起源と進化
起源
動物の起源については、単細胞生物の襟鞭毛虫が集まって多細胞化する事で海綿動物のような動物になっていったと考えられる[43]。
なお従来は、上述した襟鞭毛虫類から進化したとするヘッケルの説と繊毛虫類から進化したとするハッジの説が対立していたが、分子遺伝学の成果によれば、18S rDNAに基づいた解析等により、動物は襟鞭毛虫類を姉妹群に持つ単系統な群であることが示されており、ヘッケルの説が有力とされている[43]。
古生物
先カンブリア時代
30億年以上前に地球上初めての生物が誕生したと考えられており、真核生物の最古の化石は21億年前の地層から発見されている[44]。
動物の起源は10~12億年前まで遡れると分子系統解析と古生物学的証拠から推定されている[45]。
2012年現在、動物のものかもしれない最古の化石は2012年にナミビアの7億6000万年前の地層で発見されたオタヴィア(Otavia)という0.3~5mm程度のかりんとうのような形をした小さな殻で、海綿動物だとすると表面に空いている多数の細孔から微小なプランクトンを濾過摂食したものと考えられる[46]。またオーストラリアの南オーストラリア州からは6億6500万年のTrezona Formationという化石が発見されており、これも初期の海綿動物ではないかと考えられている[47]

エディアカラ生物群の一つであるディッキンソニア
次に古いと思われる多細胞生物の化石は6億2千万年~5億4千2百万年前のエディアカラ生物群である[48]。これらは刺胞動物、環形動物、棘皮動物の仲間であるという解釈もあれば、一部には現生動物とは全く違う動物群とする解釈もあるなど見解が定まっていない[48]。エディアカラ生物群は新原生代クライオジェニアン紀の全地球凍結(スノーボールアース)の後に進化的に拡散[訳語疑問点](Evolutionary radiation)したと考えられ(Avalon explosion、5億7500万年前)[49][50]、カンブリア爆発の頃にその多くは姿を消した(カンブリア中期の5億1000万年前~5億年前まで生き残っていたものはまれである)。
古生代

カンブリア紀の生物アノマロカリス
古生代のカンブリア紀になると、化石に残る硬い骨格を動物達が獲得し、短期間で多くのボディプランを持つ動物群が登場し[51]、海綿動物、軟体動物、腕足動物、節足動物、棘皮動物、環形動物、脊索動物など、現在の動物門のほとんどをしめる30余りの動物門が生じたとされる(カンブリア爆発)[51]。
ただし分子系統解析によればこれらの動物門は最古の化石より10億年以上遡ると推定されている[51]。カンブリア紀に突然生物が増えたように見えるのは、この時期に化石に残りやすい生物種が増えたからに過ぎない[52][53][54][55]。
カンブリア爆発は2000万年[56][57]から2500万年[58][59]続いた。オルドビス紀にはカンブリア紀までに登場した動物門が大きく適応放散している[51]。
オルドビス紀末に大量絶滅(O-S境界)があったが[51]、無顎類(顎の無い脊椎動物)は生き残り、シルル紀に多様化し、顎のある脊椎動物も登場した[51]。デボン紀には硬骨魚類が多様化し、石炭紀には両生類が繁栄、ペルム紀には爬虫類が繁栄した[51]。
シルル紀には最古の陸上動物の化石である節足動物多足類が登場し、デボン紀に節足動物が多様化、石炭紀には翅を持つ昆虫類が登場した[51]。
中生代

トリケラトプスの化石
ペルム紀末には地球史上最大の大量絶滅(P-T境界)が起こり、中生代三畳紀には海洋生物が大量に絶滅[51]。哺乳類が登場した[51]。
ジュラ紀には恐竜が繁栄し、鳥類も登場した[51]。また、軟体動物の殻を破るカニ類や硬骨魚類が進化し、これに対抗して厚い殻をもつれ合い軟体動物が進化した(中生代の海洋変革)[51]。白亜紀までには現生の昆虫類のほとんどが登場[51]。
白亜紀末には巨大隕石の衝突による大量絶滅がおこる(K-Pg境界)[51]。
新生代
新生代は哺乳類が優勢になり、鳥類、昆虫類、真骨魚類も適応放散し、現在と同様の動物相が形成される[51]。新生代の後半にあたる第四紀には人類も出現した。
化石動物についての動物門
化石動物について、上記の分類される現存動物門のいずれにも属さないとして、新たな動物門が提唱されることがある。これらについては、うたかたのごとく提唱されては消えていくものも少なくないが、主なもののみ挙げる。
- †三裂動物門 Trilobozoa Fedonkin,1985
トリブラキディウムなどが属する。三放射対称の体制をもつ。
- †盾状動物門 Proarticulata Fedonkin,1985
ディッキンソニア、ヨルギアなどのヴェンド生物が属する。左右相互に対称する体制をもつ。
- †古虫動物門 Vetulicolia Shu, et al. 2001
ウェツリコラなどが属する。その後は脊索動物とされ、その1亜門(古虫動物亜門)になる[60]。
- †葉足動物門 Lobopodia Snodgrass, 1938
アイシュアイア、ハルキゲニアなどが属する。汎節足動物であり、該当する動物門[61]のそれぞれの初期脇道系統(ステムグループ)を含んだ側系統群であると考えられる[62][63]。
絶滅した動物
現生の動物の系統
系統
この節は更新が必要とされています。 この節には古い情報が掲載されています。編集の際に新しい情報を記事に反映させてください。反映後、このタグは除去してください。(2018年7月) |
下表は動物界を生物の分類の分類階級である「門」に分類したものであり[64]、各動物門に属する生物はそれぞれの「門」独自の基本設計(ボディプラン)を共有している。
ただし、2018年現在、分子系統解析が進展中ということもあり、下表は今後も若干の修正が加えられていくものと思われる。
| 上位分類 | 門 | 既知の種の数 (2010年現在) | 動物の例 | |||
|---|---|---|---|---|---|---|
後生動物 | (前左右相称動物)[注釈 2] | 海綿動物門 | 約7000種[66] |  カイメン、 カイメン、 カイロウドウケツ カイロウドウケツ | ||
平板動物門 | 1種[67] | |||||
刺胞動物門 | 約7620種[68] |  クラゲ、 クラゲ、 サンゴ サンゴ | ||||
有櫛動物門 | 約143種[69] |  クシクラゲ クシクラゲ | ||||
左右相称動物 | 冠輪動物 | ? | 直泳動物門 | 25種[70] |  キリオキンクタ キリオキンクタ | |
| ? | 二胚動物門 (菱形動物門[70]とも) | 約110種[70] |  ニハイチュウ ニハイチュウ | |||
扁平動物 | 扁形動物門 | 約20000種[注釈 5] |  サナダムシ サナダムシ | |||
顎口動物門 | 約100種[71] | |||||
輪形動物門 | 約3000種[72] |  ワムシ ワムシ | ||||
鉤頭動物門 | 約1100種[73] |  | ||||
微顎動物門 | 1種[74] | |||||
腹毛動物門 | 約450種[75] |  イタチムシ、オビムシ イタチムシ、オビムシ | ||||
| ? | 外肛動物門 | 約4500種[76] |  コケムシ コケムシ | |||
触手冠動物 | 箒虫動物門 | 約20種[77] |  ホウキムシ ホウキムシ | |||
腕足動物門 | 約350種[77] |  シャミセンガイ、ホオズキガイ シャミセンガイ、ホオズキガイ | ||||
担輪動物 | 紐形動物門 | 約1200種[78] | ||||
軟体動物門 | 約93195種[79] |  貝類、 貝類、 イカ、 イカ、 タコ タコ | ||||
星口動物門 | 約320種[80] |  ホシムシ ホシムシ | ||||
環形動物門 | 約16650種[注釈 6] |  ミミズ、 ミミズ、 ゴカイ、 ゴカイ、 ユムシ ユムシ | ||||
内肛動物門 | 約150種[81] |  スズコケムシ スズコケムシ | ||||
有輪動物門 | 2種[82] | シンビオン | ||||
脱皮動物 | 線形動物 | 線形動物門 | 約15000種[83] |  回虫 回虫 | ||
類線形動物門 | 約320種[84] |  ハリガネムシ ハリガネムシ | ||||
有棘動物 | 動吻動物門 | 約150種[85] | トゲカワ | |||
胴甲動物門 | 約23種[86] |  コウラムシ コウラムシ | ||||
鰓曳動物門 | 約16種[87] |  エラヒキムシ エラヒキムシ | ||||
汎節足動物 | 緩歩動物門 | 約800種[88] |  クマムシ クマムシ | |||
有爪動物門 | 約160種[89] |  カギムシ カギムシ | ||||
節足動物門 | 約110万種[90] |  昆虫類、 昆虫類、 | ||||
| ? | 毛顎動物門 | 約130種[91] | ||||
新口動物 | 珍無腸動物門 | 約270種 | ||||
棘皮動物門 | 約7000種[92] |  ヒトデ、 ヒトデ、 ナマコ、 ナマコ、 ウニ ウニ | ||||
半索動物門 | 約90種[93] |  ギボシムシ、フサカツギ ギボシムシ、フサカツギ | ||||
脊索動物門 | 約51416種[94] |  ナメクジウオ、 ナメクジウオ、 ホヤ、 ホヤ、 脊椎動物 脊椎動物 | ||||

分類学の父として知られるカール・フォン・リンネ
なお、上述の分類において
- 1990年代以前は左右相称動物は原腸が口になるか否かで旧口動物、新口動物に分類され、さらに体腔が無体腔、偽体腔、真体腔のいずれであるかにより分類されていた。しかし1990年代の18S rRNA遺伝子の解析により、体腔の違いは進化とは関係ない事が判明し、上述の意味での新口動物は単系統でない事が示されたので、いくつかの動物門を新口動物から外し(後述)、全体を冠輪動物、脱皮動物、新口動物の3つに大別できることが分かった[95]。しかし以降の系統解析でも、旧口動物が単系統であること自体は支持されている[96][97][98]。
刺胞動物と有櫛動物は外形的に類似しているので腔腸動物門としてまとめられていたが、有櫛動物は刺胞がなく、上皮細胞が多繊毛性であり、決定性卵割であるといった刺胞動物との決定的違いがあり、しかも分子系統解析により腔腸動物が単系統とならないがわかったので2010年現在は両者は別の門として分けられている[99]
中生動物門は直泳動物門と二胚動物門に分けられている[64]。なお中生動物門は原生動物から後生動物に進化する過程であると過去には見られていたが、2010年現在では寄生生活により退化した後生動物であると見られている[100]。- 外肛動物は触手冠動物とされてきたが、箒虫動物や腕足動物のような他の触手冠動物どは異なる系統である事がわかった[101]。2010年現在系統上の位置は定まっていない[101]。
有髭動物門とユムシ動物門は環形動物門に入れられている[64][65]。
舌形動物門は節足動物門に入れられている[64]。- かつて扁形動物門に分類されていた珍渦虫と無腸動物については、新口動物の新たな門として珍無腸動物門が設立された[102][103]。しかし2016年の2つの分子系統解析は珍無腸動物門は左右相称動物ではあるものの、既知の3つの左右相称動物(新口動物、冠輪動物、脱皮動物)のいずれにも属さず、これら3つの姉妹群となることを支持した[104][96][105]。上の系統樹では、前者の説を前提にしたものを載せている。
前左右相称動物
前左右相称動物[注釈 2]に属する動物は体の左右相称性(=左右対称性)がない。ここに属する4つの門の間の関係、およびこれら4つの門と左右相称動物との関係は2010年現在、分子系統解析でも定まっていないる[106]。
おそらく動物全体の中で、前左右相称動物である海綿動物が最も祖先的で最初に分化した後生動物であると2017年現在考えられている[106][107][108][109][110][111][112][113] (もう一つの候補は有櫛動物[114][115][116][117] )。海綿動物は相称性がなく胚葉がないなど最も単純なボディープランを持つ[106]。海綿動物の細胞は化するものの、組織を形成することはなく[118]、複雑な器官をもたない[119]。
刺胞動物と有櫛動物の体は放射相称性を持ち、唯一の腔所である胃腔の開口は口と肛門を兼ねる[120]。これらの動物門の細胞は組織に分化しているものの、器官を形成していない[121]。中胚葉が形成されない二胚葉性の動物であるとされるが、細胞性である間充織を中胚葉とみなし、ヒドロ虫綱以外の刺胞動物と全ての有櫛動物を三胚葉性とみなす事も多い[122]。刺胞動物は手に刺胞があり、物理的または化学的刺激により毒を含む刺糸を発射する[120]。雌雄異体[120]。単体と群体がある[120]。漂泳性(クラゲ型)と付着性(ポリプ型)という生活様式の異なる2つの型を持ち[120]、両者は上下を逆さにした形[120]。それに対し有櫛動物は全てクラゲ型[123]。二放射相称で透明な脆弱な体を持つ[123]。雌雄同体[123]。
平板動物は1mm程度の薄い板上の多細胞動物で、センモウヒラムシ1種のみとされるが、様々な地域から見つかるため複数種である可能性も高い[124]。
左右相称動物
4つの門を除いた全ての動物門が左右相称動物である。左右相称動物は完全な三胚葉性で[125]、体が左右相称(=左右対称)[125]。もっとも祖先的な左右相称動物は珍無腸動物である[126][127][128]。
左右相称動物は口と肛門、およびこれらをつなぐ消化管をもち、体内に体腔ないし偽体腔(線形動物、輪形動物など)を持つ。左右相称動物のボディプランは、前方(運動のとき体の進む方向)と後方の区別、腹側と背側の区別がある傾向があり、したがって左側と右側の区別も可能である[129][130]。運動のとき体の前方へと進むので、進行方向にあるものを識別する感覚器や餌を食べる口が前方に集まる傾向にある(頭化という)。多くの左右相称動物は環状筋と縦走筋のペアを持つので[130]、ミミズのような体が柔らかい動物では水力学的骨格の蠕動により動く事ができる[131]。また多くの左右相称動物には繊毛で泳ぐことができる幼生の時期がある。
以上の特徴は例外も多い。例えば棘皮動物の成体は(幼生とは違い)放射相称であるし、寄生虫の中には極端に単純化された体の構造をもつものも多い[129][130]。
冠輪動物
巻き貝の胚の螺旋卵割
扁平動物、触手冠動物、担輪動物を含む[64]。さらに系統上の位置は不明ながら、外肛動物門も含む[64]。冠輪動物という名称は触手冠動物と担輪動物を含む事から名付けられた。なお冠輪動物(Lophotrochozoa)は狭義には扁形動物を含めず、扁形動物を含めた広義の意味で用いる場合は螺旋卵割動物(Spiralia)という名前も用いられる[132]。この名称は螺旋卵割動物のほとんど(全てではない[133])が螺旋卵割を行うという形質をもつことに注目したものである[132][134]。
扁平動物はもともとは扁形動物門、輪形動物門、鉤頭動物門に対して名付けられた門で、これら3つには体表に繊毛があり、体節がなく、循環系を欠き、無体腔または偽体腔であるという特徴がある[135]。顎口動物門、輪形動物門、鉤頭動物門の系統上の位置は2010年現在安定していないが、扁形動物の一部と近縁であると考えられている[101]。
触手冠動物は触手冠と呼ばれる独特の構造を持つ。外肛動物も同様の触手冠を持つので触手冠動物に分類されていた事もあったが、2010年現在は否定されている[136]。

受精から9時間の海洋性の腹足類Haliotis asininaのトロコフォア
担輪動物はトロコフォア型の幼生を持ち[137]、ミミズやゴカイを含む線形動物門、貝類やタコやイカを含む軟体動物門などからなる。
軟体動物門は節足動物門についで大きい門で、三胚葉性[138]。真体腔を持つ(退化的である事が多い)[138]。体節がない[138]。骨格がなく軟体[138]。一般的には体は頭部、内蔵塊、足からなり[138]、外套膜が内蔵塊を覆っている[138]。外套膜が分泌した石灰質の貝殻を持つ事が多い[138]。軟体動物門は8つの綱からなる。定説には達していないものの、8つの綱は一般的には2つの亜門に分類される[138]
- 双神経亜門:祖先的なボディープランを残している[138]
尾腔綱:蠕虫状で貝殻を持たない[138]。体長2mm~140mm程度[138]。アッケシハダウミヒモなど[138]
溝腹綱:尾腔綱と同じく蠕虫状で貝殻を持たないため、両者を合わせて無板綱と以前は呼ばれていた[138]。サンゴノフトヒモ、カセミミズなど[138]
多板綱:ヒザラガイ、オオバンヒザラガイなど[138]
- 貝類亜門:殻皮と石灰質の層状の貝殻を持つ[138]
単板綱:現生種はネオピリナなど数十種[138]。化石種は多数知られる[138]
頭足類:体が前後に伸び[138]、外套膜は内蔵塊包み胴部を形成[138]。口の周囲に触手ないし腕がある[138]。外套腔は前方に開き、腹側に筒状の漏斗を備える(水を吐いて移動したり、墨をはいたりするのに利用)[138]。貝殻はオウムガイ以外では通常体内にあるか完全に退化[138]。外套腔に鰓[138]。精巧な眼を持つ[138]。オウムガイ類、タコ類、イカ類、絶滅したアンモナイト類など[138]
腹足綱:通常は螺旋状に巻いた貝殻と蓋を持つ。カサガイ、サザエ、クロアワビ、オキナエビス、有肺類(カタツムリ、ナメクジなど)、後鰓類(ウミウシなど)を含む[138]
掘足綱:ツノガイの仲間[138]
二枚貝綱:左右に外套膜が張り出し、そこから分泌される2枚の貝殻が体の左右を覆っている[138]。ムラサキイガイ、アコヤガイ、マガキ、オカメブンブクヤドリガイ、オオシャコガイ、フナクイムシなど[138]
なお、シャミセンガイは「貝」という名称で外套膜もあるものの、触手冠動物の腕足動物門に属している。
脱皮動物

セミの脱皮
体を覆うクチクラの脱皮を行うという共通の特徴を持つ[101][139][140]。脱皮動物内の系統は2010年現在はっきりしないが、分子系統解析と形態から総合的に考えて線形動物、有棘動物、汎節足動物の3つに分類がなされている[101]。毛顎動物の系統は2010年現在不明だが、節足動物との近縁性が示唆される成果もある[101]。
線形動物はカイチュウ、ギョウチュウ、アニサキスなどからなる線形動物門とハリガネムシ目と遊線虫目からなる類線形動物門により構成される。例として寄生生物を多くあげたが、自由生活を送る線形動物もいる[141]。有棘動物は冠棘という主に頭部に数列有る環状に並ぶ棘を持つという共有性質から名付けられた[142]。花状器官という感覚器を持つことと、頭部が反転可能な事、偽体腔を持つ事も共有性質である[142][143]。
汎節足動物は、動物界最大の門である節足動物を含む系統群である。汎節足動物は体節を持つ事を特徴とする[144]。環形動物も体節を持つため、歴史的には環形動物も節足動物の近縁であると考えられていたが、分子系統解析により、近縁性が否定されたため、環形動物は汎節足動物ではなく冠輪動物に分類されている[144]。
節足動物門は以下の4つに分類される[145]。これら4つの系統的な関係性は2010年現在まだ研究中[146]。
鋏角亜門:ウミグモ類、カブトガニ類、クモ型類など
甲殻亜門:エビ・カニ類、ミジンコ類など
多足亜門:ムカデ類、ヤスデ類など
六脚亜門:広義の昆虫類。内顎類(トビムシ、カマアシムシ、コムシなど)、外顎類(狭義の昆虫類)に分かれる[147]。
汎節足動物は節足動物門以外には緩歩動物門と有爪動物門を含む。絶滅した群まで範囲を広げると葉足動物と呼ばれる古生物をも含む。緩歩動物門に属する動物はクマムシと呼ばれる動物であり[148]、ゆっくり歩く事からその名が名付けられた。陸上に生息する種では、クリプトビオシスという極限状態に耐えられる休眠状態になる事が知られている[148]。有爪動物門に属する動物はカギムシと呼ばれ、現生種は真有爪目のみ[149]。
カンブリア紀に多様化した葉足動物は、現生の汎節足動物の3つの動物門、いわゆる節足動物、緩歩動物と有爪動物のそれぞれの最後の共通祖先を含んだ側系統群であると考えられる[62][63][150]。多くの葉足動物は一見で現生の有爪動物らしい外見をもち[63]、かつては原始的な有爪動物として分類された[151][63]。しかしその後、節足動物と緩歩動物らしい形質をもつ葉足動物の発見に至り[152][153][154][155][156]、葉足動物と有爪動物の多くの共通点は、両者の系統的類縁関係を反映していない、汎節足動物の共有原始形質と見なされるようになった[150]。
新口動物

原口が口になる動物(上図)と原口が肛門になる動物(下図)。歴史的にはこれが旧口動物と新口動物を分ける基準であった。
棘皮動物門、半索動物門、脊索動物門を含み[64]、さらに珍無腸動物門も含むかもしれない(2018年現在未確定。前述)。
新口動物は歴史的には胚にできた原口が口になる旧口動物に対し、原口が口にならず新たに口が開く動物として定義されていたが[157]、1990年代に分子系統解析が始まると、この歴史的な意味での新口動物は単系統にならない事が示されたので、毛顎動物や有鬚動物などが新口動物から外され、前述の門のみが新口動物として残された[158]。この変更以降も「旧口動物」という系統群名を用いる場合もあるが[159][160][161][162][163][164][165][162] 、その場合毛顎動物や有鬚動物のような原口が口にならない動物も旧口動物に含まれている事になるので注意が必要である。なお、有鬚動物は環形動物の科にする意見が受け入れられている[166][167][168][169][170][171][172]。
脊索動物は我々脊椎動物を含む動物門で、(一生のうち少なくとも一時期に)脊索を持つという特徴をもち、以下の3つに分類される[173]:
頭索動物亜門:一生、全体長に渡って脊索を持つ。ナメクジウオの仲間
尾索動物亜門:一生ないし一時期に尾部に脊索を持つ。ホヤ綱、オタマボヤ綱、タリア綱(ヒカリボヤ、ウミタル、サルパなど)からなる。
脊椎動物亜門:脊索の周囲に脊椎が形成される。魚類、両生類、爬虫類、鳥類、哺乳類からなる
これらのうち頭索動物亜門が最も祖先的であり、尾索動物亜門と脊椎動物亜門が姉妹群をなす事が、ナメクジウオの全ゲノム解析により分かった[173]。
2010年現在、棘皮動物と半索動物が姉妹群をなすという説が大勢を締めており[174]、これら2つをあわせて歩帯動物という。歩帯動物は幼生の形態、三体腔性、軸器官などの共通性を持つ[174]。棘皮動物は、成体五放射相称[175]、三胚葉性[175]、海にのみ生息し、自由生活[175]。僅かな種をのぞき底生性[175]。雌雄異体が多い[175]。消化管は口から肛門につながるが、一部の種では肛門が退化[175]。ウミユリ綱、ヒトデ綱、クモヒトデ綱、ナマコ綱、ウニ綱からなり[175]、これらのうちウミユリ綱が最も祖先的だと考えられている[175]。半索動物はギボシムシ綱とフサカヅキ綱からなり[176]、前者は数センチから2メートル程度の蠕虫状で砂泥中に住み[176]、後者は数ミリ程度の個体が群体を作り、棲管内に住む[176]。
参考文献
- 白山義久編集;岩槻邦男・馬渡峻輔監修『無脊椎動物の多様性と系統』、(2000)、裳華房
- 藤田敏彦 『動物の系統分類と進化』 太田次郎、赤坂甲治、浅島誠、長田敏行、裳華房〈新・生命科学シリーズ〉、2010年4月28日。.mw-parser-output cite.citation{font-style:inherit}.mw-parser-output .citation q{quotes:"""""""'""'"}.mw-parser-output .citation .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .citation .cs1-lock-limited a,.mw-parser-output .citation .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .citation .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-ws-icon a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/4/4c/Wikisource-logo.svg/12px-Wikisource-logo.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-maint{display:none;color:#33aa33;margin-left:0.3em}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
ISBN 978-4785358426。 - 浅島誠、駒崎伸二 『動物の発生と分化』 太田次郎、赤坂甲治、浅島誠、長田敏行、裳華房〈新・生命科学シリーズ〉、2011年9月21日。
ISBN 978-4785358495。 - 松本忠夫 『動物の生態: 脊椎動物の進化生態を中心に』 太田次郎、赤坂甲治、浅島誠、長田敏行、裳華房〈新・生命科学シリーズ〉、2015年2月21日。
ISBN 978-4785358624。 - 馬渡, 峻輔 『動物の多様性30講』 朝倉書店〈図説生物学30講〔環境編〕3〉、2013年5月25日、初版。
ISBN 978-4-254-17723-7。
脚注
注釈
^ 左上から順に、1段目:ヒトデの一種(棘皮動物門星形動物亜門ヒトデ綱)、カイメンの一種(海綿動物門)、コウイカの一種Sepiola atlantica(軟体動物門頭足綱)、
2段目:ミズクラゲ(刺胞動物門鉢虫綱)、ガの一種Hypercompe scribonia(節足動物門六脚亜門昆虫綱)、ゴカイの一種Nereis succinea(環形動物門多毛綱)、
3段目:シャコガイの一種Tridacna squamosa(軟体動物門二枚貝綱)、シベリアトラ(脊索動物門脊椎動物亜門哺乳綱)、ホヤの一種Polycarpa aurata(脊索動物門尾索動物亜門ホヤ綱)、
4段目:クマムシの一種(緩歩動物門異クマムシ綱)、淡水産コケムシの一種(外肛動物門掩喉綱)、ウツボの一種Enchelycore anatina(脊索動物門脊椎動物亜門条鰭綱)、
5段目:カニの一種Liocarcinus vernalis(節足動物門甲殻亜門軟甲綱)、Corynosoma wegeneri(鉤頭動物門古鉤頭虫綱)、アオカケス(脊索動物門脊椎動物亜門鳥綱)、
6段目:ハエトリグモの一種(節足動物門鋏角亜門蛛形綱)、ヒラムシの一種Pseudoceros dimidiatus(扁形動物門渦虫綱)、ホウキムシ類のアクチノトロカ幼生(箒虫動物門)
- ^ abc「前左右相称動物」というのは左右相称動物以外の動物門について述べるための便宜的な名称で、「前左右相称動物」という系統群があるわけではない(藤田(2010), p.113.)
^ 両生類では内胚葉の形成と原腸の形成が同時に起こるが、鳥類や哺乳類では、内胚葉の形成がのみが原腸胚期に行われ、原腸の形成は神経胚期になってから行われる(浅島・駒崎(2011), p.126.)
^ 刺胞動物と有櫛動物が二胚葉性であるとされるが、細胞性である間充織を中胚葉とみなし、ヒドロ虫綱以外の刺胞動物と全ての有櫛動物を三胚葉性とみなす事も多い(藤田(2010) p122)
^ この数字は2010年の学術書藤田(2010), p.126によるが、扁形動物門のうち無腸動物については、別系統である事がわかり、珍無腸動物門に移された(馬渡 (2013), p27-p29)為、無腸動物の分を数字から引く必要がある。
^ 藤田(2010), pp.142-144.ではユムシ動物門約150種、環形動物門約16500種としているが、馬渡 (2013)ではユムシ動物門を環形動物門の一部としているため、合計した値をここに記した。
出典
- ^ ab“動物(どうぶつ)とは”. コトバンク. 2018年7月18日閲覧。より『デジタル大辞泉』の解説。
^ 藤田(2010) p91
^ Lisa A. Urry; Michael L. Cain; Steven A. Wasserman; Peter V. Minorsky; Jane B. Reece 池内昌彦、伊藤元己、箸本春樹 、道上達男訳 (2018/3/20). キャンベル生物学 原書11版. 丸善出版. p. 655. ISBN 978-4621302767.
^ P. レーヴン; J. ロソス; S. シンガー; G. ジョンソン (2007/5/1). レーヴン ジョンソン 生物学〈下〉(原書第7版). 培風館. p. 518.
^ “中学校理科教科書「未来へ広がるサイエンス」”. 啓林館. 2018年7月11日閲覧。
^ Avila, Vernon L. (1995). Biology: Investigating Life on Earth. Jones & Bartlett Learning. pp. 767–. ISBN 978-0-86720-942-6. https://books.google.com/books?id=B_OOazzGefEC&pg=PA767.
- ^ ab“Palaeos:Metazoa”. Palaeos. 2018年2月25日閲覧。
^ Bergman, Jennifer. “Heterotrophs”. 2007年8月29日時点のオリジナルよりアーカイブ。2007年9月30日閲覧。
^ Mentel, Marek; Martin, William (2010). “Anaerobic animals from an ancient, anoxic ecological niche”. BMC Biology 8: 32. doi:10.1186/1741-7007-8-32. PMC 2859860. PMID 20370917. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2859860/.
^ Saupe, S. G.. “Concepts of Biology”. 2007年9月30日閲覧。
^ Minkoff, Eli C. (2008). Barron's EZ-101 Study Keys Series: Biology (2nd, revised ed.). Barron's Educational Series. p. 48. ISBN 978-0-7641-3920-8.
^ 松本信二、船越浩海、玉野井逸朗 『細胞の増殖と生体システム』 学会出版センター、1993年、初版。
ISBN 4-7622-6737-6。 pp.47-50、3.細胞の微細構造とその機能、3.1.細胞と膜
^ 武村政春 『DNAを操る分子たち』 技術評論社、2012年、初版第1刷。
ISBN 978-4-7741-4998-1。 pp.14-24、第1章 エピジェネティクスを理解するための基礎知識、1-1 DNAとセントラルドグマ
^ 松本信二、船越浩海、玉野井逸朗 『細胞の増殖と生体システム』 学会出版センター、1993年、初版。
ISBN 4-7622-6737-6。、pp.53-56、3.細胞の微細構造とその機能、3.3.真核生物、3.3.1真核生物の構造と機能概説
^ 井出利憲 『細胞の運命Ⅳ細胞の老化』 サイエンス社、2006年、初版。
ISBN 4-7819-1127-7。 pp.65-75、第6章 テロメアとは何か
^ 『生化学辞典第2版』 東京化学同人、1995年、第2版第6刷。
ISBN 4-8079-0340-3。、p.534 【細胞骨格タンパク質】
^ 林純一「ミトコンドリアDNAに突然変異をもつ細胞は自然免疫により排除されることを発見」筑波大学生命科学研究科発表 「Journal of Experimental Medicine」電子版 2011.Oct.12
^ 黒岩常祥『ミトコンドリアはどこからきたか』日本放送出版、2000年6月30日第1刷発行。ISBN 4140018879。
^ Alberts, Bruce; Johnson, Alexander; Lewis, Julian; Raff, Martin; Roberts, Keith; Walter, Peter (2002). Molecular Biology of the Cell (4th ed.). Garland Science. ISBN 0-8153-3218-1. https://www.ncbi.nlm.nih.gov/books/NBK26810/.
^ Sangwal, Keshra (2007). Additives and crystallization processes: from fundamentals to applications. John Wiley and Sons. p. 212. ISBN 978-0-470-06153-4.
^ Magloire, Kim (2004). Cracking the AP Biology Exam, 2004–2005 Edition. The Princeton Review. p. 45. ISBN 978-0-375-76393-9.
^ Starr, Cecie (2007-09-25). Biology: Concepts and Applications without Physiology. Cengage Learning. pp. 362, 365. ISBN 0495381500. https://books.google.com/?id=EXNFwB-O-WUC&pg=PA362.
^ Knobil, Ernst (1998). Encyclopedia of reproduction, Volume 1. Academic Press. p. 315. ISBN 978-0-12-227020-8.
^ Schwartz, Jill (2010). Master the GED 2011. Peterson's. p. 371. ISBN 978-0-7689-2885-3.
^ Hamilton, Matthew B. (2009). Population genetics. Wiley-Blackwell. p. 55. ISBN 978-1-4051-3277-0.
- ^ abc浅島・駒崎(2011), p.33.
^ 浅島・駒崎(2011), p.45.
^ 浅島・駒崎(2011), p.39.
- ^ ab浅島・駒崎(2011), p.42.
^ 浅島・駒崎(2011), p.49.
^ 浅島・駒崎(2011), p.61.
^ 藤田(2010), p.122.
^ 藤田(2010), p.115.
^ 浅島・駒崎(2011), pp.72-73.
^ 浅島・駒崎(2011), pp72-73.
- ^ ab浅島・駒崎(2011), p.108.
- ^ abc浅島・駒崎(2011), p.115.
^ 浅島・駒崎(2011), p.110.
^ 浅島・駒崎(2011), pp.115-117.
- ^ ab浅島・駒崎(2011), p.110.
^ 浅島・駒崎(2011), p.119.
^ 浅島・駒崎(2011), p.126.
- ^ ab藤田(2010), pp.99-101.
^ 藤田(2010), p.92.
^ 藤田(2010), p.92.
^ 松本(2015) p.3.
^ Maloof, Adam C.; Rose, Catherine V.; Beach, Robert; Samuels, Bradley M.; Calmet, Claire C.; Erwin, Douglas H.; Poirier, Gerald R.; Yao, Nan et al. (17 August 2010). “Possible animal-body fossils in pre-Marinoan limestones from South Australia”. Nature Geoscience 3 (9): 653–659. Bibcode 2010NatGe...3..653M. doi:10.1038/ngeo934. http://www.nature.com/ngeo/journal/v3/n9/full/ngeo934.html.
- ^ ab藤田(2010), p.92.
^ Two Explosive Evolutionary Events Shaped Early History Of Multicellular Life
^ Shen, Bing; Dong, Lin; Xiao, Shuhai; Kowalewski, Michał (2008). “The Avalon Explosion: Evolution of Ediacara Morphospace”. Science 319 (5859): 81–84. Bibcode 2008Sci...319...81S. doi:10.1126/science.1150279. PMID 18174439. http://www.sciencemag.org/content/319/5859/81.short.
- ^ abcdefghijklmno藤田(2010), pp.92-98.
^ Maloof, A. C.; Porter, S. M.; Moore, J. L.; Dudas, F. O.; Bowring, S. A.; Higgins, J. A.; Fike, D. A.; Eddy, M. P. (2010). “The earliest Cambrian record of animals and ocean geochemical change”. Geological Society of America Bulletin 122 (11–12): 1731–1774. Bibcode 2010GSAB..122.1731M. doi:10.1130/B30346.1. http://gsabulletin.gsapubs.org/content/122/11-12/1731.
^ “New Timeline for Appearances of Skeletal Animals in Fossil Record Developed by UCSB Researchers”. The Regents of the University of California (2010年11月10日). 2014年9月1日閲覧。
^ Conway-Morris, S. (2003). “The Cambrian "explosion" of metazoans and molecular biology: would Darwin be satisfied?”. The International journal of developmental biology 47 (7–8): 505–15. PMID 14756326. http://www.ijdb.ehu.es/web/paper.php?doi=14756326.
^ “The Tree of Life”. The Burgess Shale. Royal Ontario Museum. 2018年2月28日閲覧。
^ Valentine, JW; Jablonski, D; Erwin, DH (1999). “Fossils, molecules and embryos: new perspectives on the Cambrian explosion”. Development 126 (5): 851–9. PMID 9927587. http://dev.biologists.org/content/126/5/851.long.
^ Budd, Graham (2013). “At the origin of animals: the revolutionary cambrian fossil record”. Current Genomics 14 (6): 344–354. doi:10.2174/13892029113149990011. PMC 3861885. PMID 24396267. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3861885/.
^ Erwin, D. H.; Laflamme, M.; Tweedt, S. M.; Sperling, E. A.; Pisani, D.; Peterson, K. J. (2011). “The Cambrian conundrum: early divergence and later ecological success in the early history of animals”. Science 334 (6059): 1091–1097. Bibcode 2011Sci...334.1091E. doi:10.1126/science.1206375. PMID 22116879.
^ Kouchinsky, A.; Bengtson, S.; Runnegar, B. N.; Skovsted, C. B.; Steiner, M.; Vendrasco, M. J. (2012). “Chronology of early Cambrian biomineralization”. Geological Magazine 149 (2): 221–251. Bibcode 2012GeoM..149..221K. doi:10.1017/s0016756811000720.
^ García-Bellido, Diego C; Paterson, John R (2014). “A new vetulicolian from Australia and its bearing on the chordate affinities of an enigmatic Cambrian group”. BMC Evolutionary Biology 14: 214. doi:10.1186/s12862-014-0214-z. PMC 4203957. PMID 25273382. http://www.biomedcentral.com/1471-2148/14/214/abstract#.
^ 有爪動物、緩歩動物、節足動物
- ^ abSmith, Martin R.; Ortega-Hernández, Javier (2014-08-17). “Hallucigenia’s onychophoran-like claws and the case for Tactopoda” (英語). Nature 514 (7522): 363–366. doi:10.1038/nature13576. ISSN 0028-0836. https://doi.org/10.1038/nature13576.
- ^ abcdHernández, Javier Ortega (英語). Lobopodians. http://www.academia.edu/16933971/Lobopodians.
- ^ abcdefgh藤田(2010), p.113.
- ^ ab馬渡 (2013), p.2.
^ 藤田(2010), p.115.
^ 藤田(2010), p.121.
^ 藤田(2010), p.117.
^ 藤田(2010), p.120.
- ^ abc藤田(2010), p.124.
^ 藤田(2010), p.127.
^ 藤田(2010), p.128.
^ 藤田(2010), p.129.
^ 藤田(2010), p.130.
^ 藤田(2010), p.131.
^ 藤田(2010), p.132.
- ^ ab藤田(2010), p.134.
^ 藤田(2010), p.135.
^ 藤田(2010), p.136.
^ 藤田(2010), p.142.
^ 藤田(2010), p.148.
^ 藤田(2010), p.149.
^ 藤田(2010), p.150.
^ 藤田(2010), p.152.
^ 藤田(2010), p.153.
^ 藤田(2010), p.154.
^ 藤田(2010), p.155.
^ 藤田(2010), p.156.
^ 藤田(2010), p.157.
^ 藤田(2010), p.159.
^ 藤田(2010), p.168.
^ 藤田(2010), p.169.
^ 藤田(2010), p.173.
^ 藤田(2010), p.174.
^ 藤田(2010), p.106.
- ^ abCannon, Johanna Taylor; Vellutini, Bruno Cossermelli; Smith, Julian; Ronquist, Fredrik; Jondelius, Ulf; Hejnol, Andreas (2016-02). “Xenacoelomorpha is the sister group to Nephrozoa” (英語). Nature 530 (7588): 89–93. doi:10.1038/nature16520. ISSN 0028-0836. http://www.nature.com/articles/nature16520.
^ Dunn, Casey W.; Hejnol, Andreas; Matus, David Q.; Pang, Kevin; Browne, William E.; Smith, Stephen A.; Seaver, Elaine; Rouse, Greg W. et al. (2008-03-05). “Broad phylogenomic sampling improves resolution of the animal tree of life” (英語). Nature 452 (7188): 745–749. doi:10.1038/nature06614. ISSN 0028-0836. http://www.nature.com/doifinder/10.1038/nature06614.
^ Egger, Bernhard; Steinke, Dirk; Tarui, Hiroshi; Mulder, Katrien De; Arendt, Detlev; Borgonie, Gaëtan; Funayama, Noriko; Gschwentner, Robert et al. (2009-05-11). “To Be or Not to Be a Flatworm: The Acoel Controversy” (英語). PLOS ONE 4 (5): e5502. doi:10.1371/journal.pone.0005502. ISSN 1932-6203. PMC PMC2676513. PMID 19430533. http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0005502.
^ 藤田(2010), p.119.
^ 藤田(2010), p.122.
- ^ abcdef藤田(2010), pp.122-123.
^ Philippe, Hervé; Brinkmann, Henner; Copley, Richard R.; Moroz, Leonid L.; Nakano, Hiroaki; Poustka, Albert J.; Wallberg, Andreas; Peterson, Kevin J. et al. (2011-02). “Acoelomorph flatworms are deuterostomes related to Xenoturbella” (英語). Nature 470 (7333): 255–258. doi:10.1038/nature09676. ISSN 0028-0836. PMC PMC4025995. PMID 21307940. http://www.nature.com/articles/nature09676.
^ 馬渡 (2013), p27-p29
^ Rouse, Greg W.; Wilson, Nerida G.; Carvajal, Jose I.; Vrijenhoek, Robert C. (2016-02). “New deep-sea species of Xenoturbella and the position of Xenacoelomorpha” (英語). Nature 530 (7588): 94–97. doi:10.1038/nature16545. ISSN 0028-0836. http://www.nature.com/articles/nature16545.
^ “分類学:珍無腸動物門はNephrozoaの姉妹群である”. ネイチャー (2016年2月4日). 2018年7月20日閲覧。
- ^ abc藤田(2010), p.114.
^ Bhamrah, H. S.; Juneja, Kavita (2003). An Introduction to Porifera. Anmol Publications. p. 58. ISBN 978-81-261-0675-2.
^ Feuda, Roberto; Dohrmann, Martin; Pett, Walker; Philippe, Hervé; Rota-Stabelli, Omar; Lartillot, Nicolas; Wörheide, Gert; Pisani, Davide (2017). “Improved Modeling of Compositional Heterogeneity Supports Sponges as Sister to All Other Animals”. Current Biology 27 (24): 3864. doi:10.1016/j.cub.2017.11.008. PMID 29199080. http://linkinghub.elsevier.com/retrieve/pii/S0960982217314537.
^ Pisani, Davide; Pett, Walker; Dohrmann, Martin; Feuda, Roberto; Rota-Stabelli, Omar; Philippe, Hervé; Lartillot, Nicolas; Wörheide, Gert (15 December 2015). “Genomic data do not support comb jellies as the sister group to all other animals”. Proceedings of the National Academy of Sciences 112 (50): 15402–15407. Bibcode 2015PNAS..11215402P. doi:10.1073/pnas.1518127112. PMC 4687580. PMID 26621703. http://www.pnas.org/content/112/50/15402.
^ Simion, Paul; Philippe, Hervé; Baurain, Denis; Jager, Muriel; Richter, Daniel J.; Franco, Arnaud Di; Roure, Béatrice; Satoh, Nori et al. (3 April 2017). “A Large and Consistent Phylogenomic Dataset Supports Sponges as the Sister Group to All Other Animals”. Current Biology 27 (7): 958–967. doi:10.1016/j.cub.2017.02.031. PMID 28318975. https://doi.org/10.1016/j.cub.2017.02.031.
^ Giribet, Gonzalo (1 October 2016). “Genomics and the animal tree of life: conflicts and future prospects”. Zoologica Scripta 45: 14–21. doi:10.1111/zsc.12215. http://onlinelibrary.wiley.com/doi/10.1111/zsc.12215/abstract.
^ Laumer, Christopher E.; Gruber-Vodicka, Harald; Hadfield, Michael G.; Pearse, Vicki B.; Riesgo, Ana; Marioni, John C.; Giribet, Gonzalo (2017). “Placozoans are eumetazoans related to Cnidaria”. BioRxiv: 200972. doi:10.1101/200972. https://www.biorxiv.org/content/early/2017/10/11/200972.
^ Eitel, Michael; Francis, Warren; Osigus, Hans-Jürgen; Krebs, Stefan; Vargas, Sergio; Blum, Helmut; Williams, Gray Argust; Schierwater, Bernd et al. (2017-10-13). “A taxogenomics approach uncovers a new genus in the phylum Placozoa”. BioRxiv: 202119. doi:10.1101/202119. https://www.biorxiv.org/content/early/2017/10/13/202119.
^ Dunn, Casey W.; Hejnol, Andreas; Matus, David Q.; Pang, Kevin; Browne, William E.; Smith, Stephen A.; Seaver, Elaine; Rouse, Greg W. et al. (2008). “Broad phylogenomic sampling improves resolution of the animal tree of life”. Nature 452 (7188): 745–749. Bibcode 2008Natur.452..745D. doi:10.1038/nature06614. PMID 18322464. http://www.nature.com/doifinder/10.1038/nature06614.
^ Whelan, Nathan V.; Kocot, Kevin M.; Moroz, Tatiana P.; Mukherjee, Krishanu; Williams, Peter; Paulay, Gustav; Moroz, Leonid L.; Halanych, Kenneth M. (2017). “Ctenophore relationships and their placement as the sister group to all other animals”. Nature Ecology & Evolution 1 (11): 1737. doi:10.1038/s41559-017-0331-3. http://www.nature.com/articles/s41559-017-0331-3.
^ Shen, Xing-Xing; Hittinger, Chris Todd; Rokas, Antonis (2017-04-10). “Contentious relationships in phylogenomic studies can be driven by a handful of genes”. Nature Ecology & Evolution 1 (5): 0126. doi:10.1038/s41559-017-0126. http://www.nature.com/articles/s41559-017-0126.
^ Ryan, Joseph F.; Pang, Kevin; Schnitzler, Christine E.; Nguyen, Anh-Dao; Moreland, R. Travis; Simmons, David K.; Koch, Bernard J.; Francis, Warren R. et al. (13 December 2013). “The Genome of the Ctenophore Mnemiopsis leidyi and Its Implications for Cell Type Evolution”. Science 342 (6164): 1242592. doi:10.1126/science.1242592. PMC 3920664. PMID 24337300. http://science.sciencemag.org/content/342/6164/1242592.
^ Jessop, Nancy Meyer (1970). Biosphere; a study of life. Prentice-Hall. p. 428.
^ Sumich, James L. (2008). Laboratory and Field Investigations in Marine Life. Jones & Bartlett Learning. p. 67. ISBN 978-0-7637-5730-4.
- ^ abcdef藤田(2010), pp.117-120.
^ Safra, Jacob E. (2003). The New Encyclopædia Britannica, Volume 16. Encyclopædia Britannica. p. 523. ISBN 978-0-85229-961-6.
^ 藤田(2010) p122
- ^ abc藤田(2010), pp.120-121.
^ 藤田(2010), p.122.
- ^ ab藤田(2010), pp.122-132
^ Philippe, H.; Brinkmann, H.; Copley, R. R.; Moroz, L. L.; Nakano, H.; Poustka, A. J.; Wallberg, A.; Peterson, K. J. et al. (2011). “Acoelomorph flatworms are deuterostomes related to Xenoturbella”. Nature 470 (7333): 255–258. Bibcode 2011Natur.470..255P. doi:10.1038/nature09676. PMC 4025995. PMID 21307940. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4025995/.
^ Perseke, M.; Hankeln, T.; Weich, B.; Fritzsch, G.; Stadler, P.F.; Israelsson, O.; Bernhard, D.; Schlegel, M. (August 2007). “The mitochondrial DNA of Xenoturbella bocki: genomic architecture and phylogenetic analysis”. Theory Biosci 126 (1): 35–42. doi:10.1007/s12064-007-0007-7. PMID 18087755. http://www.bioinf.uni-leipzig.de/Publications/PREPRINTS/07-009.pdf.
^ Cannon, Johanna T.; Vellutini, Bruno C.; Smith III, Julian.; Ronquist, Frederik; Jondelius, Ulf; Hejnol, Andreas (3 February 2016). “Xenacoelomorpha is the sister group to Nephrozoa”. Nature 530 (7588): 89–93. Bibcode 2016Natur.530...89C. doi:10.1038/nature16520. PMID 26842059. http://www.nature.com/nature/journal/v530/n7588/full/nature16520.html 2016年2月3日閲覧。.
- ^ abMinelli, Alessandro (2009). Perspectives in Animal Phylogeny and Evolution. Oxford University Press. p. 53. ISBN 978-0-19-856620-5. https://books.google.com/books?id=jIASDAAAQBAJ&pg=PA53.
- ^ abcBrusca, Richard C. (2016). Introduction to the Bilateria and the Phylum Xenacoelomorpha | Triploblasty and Bilateral Symmetry Provide New Avenues for Animal Radiation. Sinauer Associates. pp. 345–372. ISBN 978-1605353753. http://www.sinauer.com/media/wysiwyg/samples/Brusca3e_Chapter_9.pdf.
^ Quillin, K. J. (May 1998). “Ontogenetic scaling of hydrostatic skeletons: geometric, static stress and dynamic stress scaling of the earthworm lumbricus terrestris”. The Journal of Experimental Biology 201 (12): 1871–83. PMID 9600869. http://jeb.biologists.org/cgi/pmidlookup?view=long&pmid=9600869.
- ^ abStruck, T. H.; Wey-Fabrizius, A. R.; Golombek, A.; Hering, L.; Weigert, A.; Bleidorn, C.; Klebow, S.; Iakovenko, N. et al. (2014), “Platyzoan paraphyly based on phylogenomic data supports a noncoelomate ancestry of Spiralia”, Molecular Biology and Evolution 31 (7): 1833–1849, doi:10.1093/molbev/msu143
^ “Explanations.html”. 2009年6月28日閲覧。
^ Shankland, M.; Seaver, E. C. (2000). “Evolution of the bilaterian body plan: What have we learned from annelids?”. Proceedings of the National Academy of Sciences 97 (9): 4434–7. Bibcode 2000PNAS...97.4434S. doi:10.1073/pnas.97.9.4434. JSTOR 122407. PMC 34316. PMID 10781038. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC34316/.
^ 藤田(2010), p.126.
^ 藤田(2010), pp.132
^ 藤田(2010), pp.135
- ^ abcdefghijklmnopqrstuvwxyzaaabacad藤田(2010), pp.136-137.
^ 藤田(2010), pp.108
^ Dawkins, Richard (2005). The Ancestor's Tale: A Pilgrimage to the Dawn of Evolution. Houghton Mifflin Harcourt. p. 381. ISBN 978-0-618-61916-0.
^ 藤田(2010), pp.150-152
- ^ ab藤田(2010), pp.153
^ Miller, Stephen A.; Harley, John P. (2006). Zoology. McGraw-Hill Higher Education. p. 173. https://books.google.com/books?id=BWZFAQAAIAAJ.
- ^ ab藤田(2010), pp.155-156
^ 藤田(2010), p.159.
^ 藤田(2010), p.160.
^ 藤田(2010), p.168.
- ^ ab藤田(2010), p.157.
^ 藤田(2010), pp.157-158.
- ^ abSiveter, Derek J.; Briggs, Derek E. G.; Siveter, David J.; Sutton, Mark D.; Legg, David (2018-08-01). “A three-dimensionally preserved lobopodian from the Herefordshire (Silurian) Lagerstätte, UK” (英語). Open Science 5 (8): 172101. doi:10.1098/rsos.172101. ISSN 2054-5703. http://rsos.royalsocietypublishing.org/content/5/8/172101.
^ Ramsköld, L.; Xianguang, Hou (1991-05). “New early Cambrian animal and onychophoran affinities of enigmatic metazoans” (英語). Nature 351 (6323): 225–228. doi:10.1038/351225a0. ISSN 0028-0836. https://www.nature.com/articles/351225a0.
^ Budd, Graham (1993-08). “A Cambrian gilled lobopod from Greenland” (英語). Nature 364 (6439): 709–711. doi:10.1038/364709a0. ISSN 0028-0836. https://doi.org/10.1038/364709a0.
^ BUDD, GRAHAM E. (1996-03). “The morphology of Opabinia regalis and the reconstruction of the arthropod stem-group” (英語). Lethaia 29 (1): 1–14. doi:10.1111/j.1502-3931.1996.tb01831.x. ISSN 0024-1164. https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1502-3931.1996.tb01831.x.
^ “(PDF) Morpho-anatomy of the lobopod Magadictyon cf. haikouensis from the Early Cambrian Chengjiang Lagerstätte, South China (Acta Zoologica (2007) DOI: 10.1111/j.1463-6395.2007.00281.x)” (英語). ResearchGate. 2018年10月28日閲覧。
^ Dzik, Jerzy (2011). "The xenusian-to-anomalocaridid transition within the lobopodians". Bollettino della Società Paleontologica Italiana, 50(1): 65-74.
^ Budd, Graham E. (2001-01). “Tardigrades as ‘Stem-Group Arthropods’: The Evidence from the Cambrian Fauna”. Zoologischer Anzeiger - A Journal of Comparative Zoology 240 (3-4): 265–279. doi:10.1078/0044-5231-00034. ISSN 0044-5231. https://doi.org/10.1078/0044-5231-00034.
^ 藤田(2010) p104
^ 藤田(2010) p108
^ Edgecombe, Gregory D.; Giribet, Gonzalo; Dunn, Casey W.; Hejnol, Andreas; Kristensen, Reinhardt M.; Neves, Ricardo C.; Rouse, Greg W.; Worsaae, Katrine et al. (June 2011). “Higher-level metazoan relationships: recent progress and remaining questions”. Organisms, Diversity & Evolution 11 (2): 151–172. doi:10.1007/s13127-011-0044-4.
^ Fröbius, Andreas C.; Funch, Peter (2017-04-04). “Rotiferan Hox genes give new insights into the evolution of metazoan bodyplans”. Nature Communications 8 (1). Bibcode 2017NatCo...8....9F. doi:10.1038/s41467-017-00020-w. http://www.nature.com/articles/s41467-017-00020-w.
^ Smith, Martin R.; Ortega-Hernández, Javier (2014). “Hallucigenia’s onychophoran-like claws and the case for Tactopoda”. Nature 514 (7522): 363–366. Bibcode 2014Natur.514..363S. doi:10.1038/nature13576. https://doi.org/10.1038/nature13576.
- ^ ab“Palaeos Metazoa: Ecdysozoa”. palaeos.com. 2017年9月2日閲覧。
^ Yamasaki, Hiroshi; Fujimoto, Shinta; Miyazaki, Katsumi (June 2015). “Phylogenetic position of Loricifera inferred from nearly complete 18S and 28S rRNA gene sequences”. Zoological Letters 1: 18. doi:10.1186/s40851-015-0017-0. https://doi.org/10.1186/s40851-015-0017-0.
^ Nielsen, C. (2002). Animal Evolution: Interrelationships of the Living Phyla (2nd ed.). Oxford University Press. ISBN 0-19-850682-1.
^ “Bilateria”. Tree of Life Web Project (2001年). 2014年8月11日閲覧。
^ 三浦「有鬚動物」『無脊椎動物の多様性と系統』(2000)
^ 小島「ハオリムシ類の進化と系統」(2008) および三浦・藤倉 (2008)
^ 三浦「環形動物」『動物系統分類学追補版』(2000)
^ 「有鬚動物門」『図説 無脊椎動物学』(2009、原書は2001)
^ Pleijel et al. (2009)
^ WoRMS (2010年). “Pogonophora”. World Register of Marine Species. 2011年6月18日閲覧。
^ Shapiro L.. “Pogonophora: Comprehensive Description”. Encyclopedia of Life. 2011年6月18日閲覧。
- ^ ab藤田(2010), pp.174-175.
- ^ ab藤田(2010), p.169.
- ^ abcdefgh藤田(2010), pp.169-172.
- ^ abc藤田(2010), p.173.
関連項目
- 動物園
- 獣医師
- 動物の行動
.mw-parser-output .hlist ul,.mw-parser-output .hlist ol{padding-left:0}.mw-parser-output .hlist li,.mw-parser-output .hlist dd,.mw-parser-output .hlist dt{margin-right:0;display:inline-block}.mw-parser-output .hlist dt:after,.mw-parser-output .hlist dd:after,.mw-parser-output .hlist li:after{white-space:normal}.mw-parser-output .hlist li:after,.mw-parser-output .hlist dd:after{content:" · ";font-weight:bold}.mw-parser-output .hlist dt:after{content:": "}.mw-parser-output .hlist-pipe dd:after,.mw-parser-output .hlist-pipe li:after{content:" | ";font-weight:normal}.mw-parser-output .hlist-hyphen dd:after,.mw-parser-output .hlist-hyphen li:after{content:" - ";font-weight:normal}.mw-parser-output .hlist-comma dd:after,.mw-parser-output .hlist-comma li:after{content:"、 ";font-weight:normal}.mw-parser-output .hlist dd:last-child:after,.mw-parser-output .hlist dt:last-child:after,.mw-parser-output .hlist li:last-child:after{content:none}.mw-parser-output .hlist dd dd:first-child:before,.mw-parser-output .hlist dd dt:first-child:before,.mw-parser-output .hlist dd li:first-child:before,.mw-parser-output .hlist dt dd:first-child:before,.mw-parser-output .hlist dt dt:first-child:before,.mw-parser-output .hlist dt li:first-child:before,.mw-parser-output .hlist li dd:first-child:before,.mw-parser-output .hlist li dt:first-child:before,.mw-parser-output .hlist li li:first-child:before{content:" (";font-weight:normal}.mw-parser-output .hlist dd dd:last-child:after,.mw-parser-output .hlist dd dt:last-child:after,.mw-parser-output .hlist dd li:last-child:after,.mw-parser-output .hlist dt dd:last-child:after,.mw-parser-output .hlist dt dt:last-child:after,.mw-parser-output .hlist dt li:last-child:after,.mw-parser-output .hlist li dd:last-child:after,.mw-parser-output .hlist li dt:last-child:after,.mw-parser-output .hlist li li:last-child:after{content:") ";font-weight:normal}.mw-parser-output .hlist ol{counter-reset:listitem}.mw-parser-output .hlist ol>li{counter-increment:listitem}.mw-parser-output .hlist ol>li:before{content:" "counter(listitem)" ";white-space:nowrap}.mw-parser-output .hlist dd ol>li:first-child:before,.mw-parser-output .hlist dt ol>li:first-child:before,.mw-parser-output .hlist li ol>li:first-child:before{content:" ("counter(listitem)" "}
- 本能
- 反射
- 走性
学習(刷り込み)- 条件反射
- 知能
畜生 - 仏教において動物を意味する
| ||||||||||||


